

. 24. júla 2023; 11:151. doi: 10.1186/s40168-023-01599-7
Abstrakt
Pozadie
Kliešte môžu prenášať širokú škálu medicínsky významných patogénov vrátane Borrelia afzelii , pôvodcu lymskej boreliózy v Európe. Kliešťová mikroflóra je dôležitým faktorom modulujúcim nielen fyziológiu vektorov, ale aj kompetenciu vektorov. Antimikrobiota vakcíny zamerané na kľúčové taxóny kliešťovej mikrobioty môžu zmeniť kŕmenie kliešťom a modulovať taxonomické a funkčné profily bakteriálnych spoločenstiev vo vektore. Vplyv antimikrobiotickej vakcíny na vývoj patogénu prenášaného kliešťami v rámci vektora však nebol testovaný.
Výsledky
Tu sme charakterizovali moduláciu mikrobioty Ixodes ricinus v reakcii na infekciu B. afzelii a zistili sme, že patogén vyvoláva zmeny v zložení mikrobioty, jej beta diverzite a štruktúre zostavy bakteriálnej komunity. Perturbácia kliešťovej mikroflóry protilátkami proti mikroflóre alebo pridanie nových komenzálnych baktérií do stredného čreva kliešťa spôsobuje odchýlky od B. afzelii -indukovanej modulácie kliešťovej mikroflóry, čo viedlo k nižšej záťaži patogénom v I. ricinus . Siete spoločného výskytu umožnili identifikáciu vznikajúcich vlastností bakteriálnych komunít, ktoré lepšie definovali stavy mikrobioty kliešťov odolné voči infekcii Borrelia .
Závery
Tieto zistenia naznačujú, že Borrelia je vysoko citlivá na poruchy kliešťovej mikrobioty a že odklon od modulácie indukovanej patogénom vo vektorovej mikrobiote predstavuje pre spirochétu vysoké náklady. Sieťová analýza sa javí ako vhodný nástroj na identifikáciu vznikajúcich vlastností vektorovej mikrobioty spojenej so stavmi odolnými voči infekcii. Antimikrobiálne vakcíny sa môžu použiť ako nástroj na narušenie mikroflóry a kontrolu dôležitých patogénov prenášaných vektormi.
Doplňujúce informácie
Online verzia obsahuje doplnkový materiál dostupný na 10.1186/s40168-023-01599-7.
Pozadie
Kliešte sú prenášačmi množstva patogénov, ktoré môžu spôsobiť infekčné ochorenia medicínskeho a veterinárneho významu. Jedným z hlavných príkladov choroby prenášanej kliešťami je lymská borelióza [ 1 ], ktorá je spôsobená genodruhovým komplexom spirochéty Borrelia burgdorferi sensu lato (sl) [ 2 ]. Spomedzi genodruhov je pôvodcom väčšiny prípadov lymskej boreliózy v Európe Borrelia afzelii [ 3 ]. Borélie sa v prírode udržiavajú vďaka biologickému prenosu sprostredkovanému kliešťami rodu Ixodes [ 4 ]. Spirochéty borélií sa vo všeobecnosti získavajú larválnymi alebo nymfálnymi štádiami kliešťov, ktoré sa živia krvou infikovaného hostiteľa stavovcov [ 5 ]. Po požití kliešťami sa spirochéty dostanú do čreva a kolonizujú ho. Po línaní kliešťa, pri ďalšom krvnom jedle, migrujú borélie z čreva do slinnej žľazy a sú prenášané nymfami alebo dospelými jedincami na nového hostiteľa spolu so slinami kliešťa [ 2 , 3 ]. V tomto cykle získavania, kolonizácie a prenosu spirochét Borrelia kliešťami môže na ňom zohrávať kľúčovú úlohu skupina endogénnych baktérií, ktoré tvoria mikrobiotu kliešťov.
Niekoľko štúdií preukázalo, že mikrobiota môže formovať kompetenciu vektora pre patogény u rôznych článkonožcov [ 6 – 10 ]. Napríklad u kliešťov narušenie mikroflóry Dermacentor andersoni , kliešťa z Rocky Mountain na báze antibiotík, znížilo získanie patogénu Francisella novicida [ 11 ]. Okrem toho hladina F. novicida pozitívne korelovala so znížením množstva endosymbiontov Francisella v mikrobiote D. andersoni, čo dokazuje pozitívny vzťah patogén-endosymbiont [ 11 ]. Perturbácia Ixodes scapularis microbiota do dysbiózneho stavu znížila kolonizáciu B. burgdorferi sensu stricto u lariev [ 9 ]. Abundancia B. burgdorferi v kliešťoch I. scapularis negatívne korelovala s abundanciou niektorých bakteriálnych taxónov, ako sú Pseudomonas alebo Staphylococcus , a pozitívne korelovala so Sphingomonas [ 12 ]. Asociácie medzi komenzálnymi baktériami a hladinami patogénov v kliešťoch naznačujú intímne interakcie patogén-mikrobiota, ktoré by mohli uľahčiť alebo obmedziť kolonizáciu patogénom vo vektore. Zacielenie na špecifické baktérie vektorovej mikrobioty, ktoré uľahčujú kolonizáciu patogénom, by mohlo byť možnou metódou kontroly prostredníctvom vakcín blokujúcich prenos.
Vo všeobecnosti sa experimentálna manipulácia s mikroflórou dosiahla expozíciou antibiotika alebo sterilnými podmienkami chovu vektora. Tieto metódy však vyvolávajú globálne zmeny v mikrobiote a sťažujú vyčerpanie špecifických baktérií. Nedávno boli navrhnuté antimikrobiálne vakcíny ako presný nástroj na manipuláciu s mikrobiotami [ 13 , 14 ]. Najmä identifikácia kľúčového taxónu (tj vysoko prepojených taxónov, ktoré riadia zloženie a funkciu komunity), Enterobacteriaceae a následná vakcinácia proti nemu vyvolali hostiteľské protilátky, ktoré boli požité vektorom počas krvného jedla a korelovali so zníženým výskytom Enterobacteriaceae v Ixodes ricinus microbiota [ 14 ]. Okrem toho antimikrobiota vakcína ovplyvnila fyziológiu kliešťov zvýšením hmotnosti kliešťa počas kŕmenia [ 13 ] a modulovala zloženie a diverzitu mikrobioty kliešťa spôsobom špecifickým pre taxón [ 14 ]. Vplyv antimikrobiálnych vakcín na vývoj patogénov bol preukázaný u Plasmodium relictum a vektora komára Culex quinquefasciatus [ 15 ]. Imunitné zacielenie na vektor-asociované Enterobacteriaceae modulovalo zloženie a diverzitu mikrobioty C. quinquefasciatus a znížilo výskyt a abundanciu P. relictum v stredných črevách a slinných žľazách komárov [ 15 ].
V tejto štúdii sa zameriavame na testovanie, či manipulácia s kliešťovou mikroflórou vakcináciou hostiteľských myší antimikrobiotami proti kľúčovému taxónu Enterobacteriaceae znižuje kolonizáciu B. afzelii vo vektore I. ricinus . Porovnanie neinfikovanej kliešťovej mikroflóry s mikroflórou vystavenou infekcii B. afzelii , antimikrobiálnymi protilátkami a novou komenzálnou baktériou umožnilo identifikáciu infekčne permisívnych a na infekciu odolných stavov mikrobiálnych spoločenstiev. Výsledky budú informovať o nových intervenciách na kontrolu lymskej boreliózy a iných chorôb prenášaných vektormi.
Materiály a metódy
Etické vyhlásenie
Experimenty in vivo sa uskutočnili v zariadení pre zvieratá Laboratória pre zdravie zvierat Francúzskej agentúry pre bezpečnosť potravín, životného prostredia a pri práci (ANSES), Maisons-Alfort, Francúzsko, podľa francúzskych a medzinárodných hlavných zásad pre biomedicínsky výskum zahŕňajúci zvieratá (2012). Postupy preverila a schválila Etická komisia (ComEth, Anses/ENVA/UPEC) s číslom povolenia na pokusy na zvieratách E 94 046 08.
Myši a podmienky bývania
Šesťtýždňové samice myší C3H/HeN (kód kmeňa Charles River 025) boli zakúpené od Charles River (Miserey, Francúzsko) a uchovávané na adaptáciu 1 týždeň pred uskutočnením experimentov. Počas štúdie boli myši chované vo vetraných stojanoch so zelenou linkou (Tecniplast, Hohenpeissenberg, Nemecko) pri -20 Pa, s jedlom (Kliba nafaj, Rinaustrasse, Švajčiarsko) a vodou ad libitum. Myši sa udržiavali pri kontrolovanej izbovej teplote (RT, 20–23 ° C) a režime fotoperiód 12 hodín (h) svetlo: 12 hodín tma. Počet myší na klietku bol obmedzený na päť. Zvieratá boli monitorované dvakrát denne (d) skúsenými technikmi a odchýlky od normálneho správania alebo známky zhoršenia zdravia boli zaznamenané a zaznamenané.
Bakteriálne kultúry
B. afzelii CB43 s nízkou pasážou sa začali z glycerolových zásob a pestovali sa v médiu Barbour-Stoenner-Kelly (BSK)-H (Sigma-Aldrich, St. Louis, MO, USA) obsahujúcom 6 % králičieho séra a udržiavali sa pri teplote 33 °C počas 7 dní. Escherichia coli BL21 (DE3, Invitrogen, Carlsbad, CA, USA) sa pestovala v Lysogeny Broth (LB, Sigma-Aldrich, St. Louis, MO, USA) pri 37 °C za intenzívneho miešania cez noc.
Experimentálna infekcia myší B. afzelii
Pri infekcii boréliami sa 1 x 106 B. afzelii CB43 v 250 ul média BSK-H injikovalo subkutánne (100 ul) a intraperitoneálne (150 ul) myšiam C3H/HeN. Kontrolným myšiam sa injikovalo samotné médium BSK-H podľa rovnakého protokolu, ako je opísané vyššie. Vzorky krvi boli odobraté zvieratám zo všetkých experimentálnych skupín 3 týždne po inokulácii na potvrdenie infekcie pomocou Western blotu [ 16 ]. Okrem toho sa od každej myši zo všetkých experimentálnych skupín na konci experimentu odobral pravý členkový kĺb, srdce a koža, aby sa potvrdila infekcia pomocou qPCR (pozri nižšie).
Očkovanie živými baktériami
Živá bakteriálna vakcína bola pripravená s použitím E. coli BL21 (DE3, Invitrogen, Carlsbad, CA, USA), ako už bolo opísané [ 13 ]. Stručne, kultúra E. coli sa premyla fyziologickým roztokom tlmivého fosforečnanu (PBS) 10 mM NaH2P04, 2,68 mM KCI, 140 mM NaCl, pH 7,2 (Thermo Scientific, Waltham, MA, USA), resuspendovala sa pri 3,6 x 104 CFU (kolóniách) a homogenizovala sa sklenená jednotka na homogenizáciu/ml. C3H/HeN myši boli imunizované subkutánne 100 ul E. coli BL21 (1 x 106 CFU na myš) v emulzii vody v oleji obsahujúcej 70 % adjuvans Montanide™ ISA 71 VG (Seppic, Paríž, Francúzsko), s posilňovacou dávkou 2 týždne po prvej dávke. Kontrolné myši dostali falošnú vakcínu obsahujúcu PBS a adjuvans.
Napadnutie kliešťami
Nekŕmené larvy I. ricinus bez patogénov sa získali z kolónií UMR BIPAR, Maisons-Alfort, Francúzsko. Myši boli anestetizované izofluránom a kapsula z EVA peny s vonkajším priemerom 2 cm (Cosplay Shop, Brugge, Belgicko) bola prilepená na ich oholený chrbát pomocou nedráždivého latexového lepidla (Tear Mender, USA), ako je opísané v Mateos-Hernandez et al. [ 17 ]. Každá myš v rôznych skupinách bola zamorená stovkou lariev I. ricinus na 30. deň (obr. 1 ). Kliešte umiestnené v injekčnej striekačke boli vložené do kapsuly pomalým zatlačením piestu a potom bolo na uzavretie kapsuly použité plastové viečko [ 17 ]. Kŕmenie kliešťov bolo vizuálne monitorované dvakrát denne. Prekrvené larvy sa odobrali do sterilných skúmaviek s otvormi a udržiavali sa s cyklom svetlo-tma (12 h/12 h) v inkubátore s relatívnou vlhkosťou > 97 % pri 22 °C.
Obr.

Príprava vzorky séra
Vzorky krvi získané retroorbitálnym krvácaním sa odobrali do sterilných skúmaviek v deň 0 a deň 30 zvieratám zo všetkých experimentálnych skupín. Okrem toho sa vzorky krvi odobrali na 14., 45. a 52. deň myšiam zo skupín PBS + B. afzelii a E. coli + B. afzelii . Vzorky krvi sa inkubovali počas 2 hodín pri teplote miestnosti bez antikoagulantu, čo umožnilo zrážanie, a potom sa dvakrát centrifugovali pri 5000 x g počas 5 minút pri teplote miestnosti. Séra sa potom oddelili a skladovali v nových sterilných skúmavkách pri teplote -20 °C až do použitia.
Bakteriálna extrakcia proteínov
Lyzáty kultúry B. afzelii boli pripravené na uskutočnenie Western blotov. Sedem mililitrov kultúry B. afzelii s hustotou aspoň 1 x 107 /ml sa centrifugovalo pri 8000 ot./min. počas 10 minút pri 20 °C. Supernatant sa potom odstránil a bakteriálna peleta sa dvakrát premyla 1 ml studeného HN-tlmivého roztoku, centrifugovala sa pri 8000 ot./min. počas 10 minút pri 20 °C. Výsledná peleta bola resuspendovaná v 200 ul bakteriálneho proteínového extrakčného (B-PER) pufra (Thermo Scientific, Waltham, MA, USA) a inkubovaná pri RT počas 10 minút. Lyzát sa skladoval pri -20 °C až do použitia. Uskutočnila sa extrakcia proteínu Escherichia coli a neskôr sa použila na potiahnutie antigénom pre test ELISA. Kultúra E. coli bola dvakrát premytá PBS, centrifugovaná pri 1000 x g počas 5 minút pri 4 °C, resuspendovaná v 1% Triton-PBS lyzovacom pufri (Sigma-Aldrich, St. Louis, MO, USA) a homogenizovaná 20 údermi s použitím homogenizátora so sklenenými guľôčkami. Homogenát sa potom centrifugoval pri 300 x g počas 5 minút pri 4 ° C a supernatant sa zhromaždil. Koncentrácia proteínov B. afzelii a E. coli bola stanovená pomocou Bradfordovho proteínového testu (Thermo Scientific, San Jose, CA, USA) s hovädzím sérovým albumínom (BSA) ako štandardom.
Western blot
Infekčnosť B. afzelii u infikovaných myší bola kontrolovaná westernovým prenosom s použitím séra myší ako primárnych protilátok. Lyzáty B. afzelii sa zmiešali s rovnakým objemom 2X Laemmliho pufra (Thermo Scientific, Waltham, Massachusetts, USA) na konečné množstvo 20 ug proteínu/pás a denaturovali sa teplom pri 100 °C počas 10 minút. Pripravené lyzáty sa naniesli do 4–15% proteínového gélu Mini-PROTEAN TGX Stain-Free (Bio-Rad, Hercules, CA, USA) a potom sa uskutočnila elektroforéza SDS-page pri 120 V počas 1 hodiny. Proteíny sa potom preniesli na nitrocelulózovú membránu (Bio-Rad, Hercules, CA, USA) s použitím metódy polosuchého prenosu. Prenos sa uskutočňoval počas 30 minút pri 25 V v prenosovej cele (Trans-Blot SD, Bio-Rad, Hercules, CA, USA). Imunoblotovanie sa uskutočnilo blokovaním membrány 1% hovädzím sérovým albumínom (BSA)/PBS (Sigma-Aldrich, St. Louis, MO, USA) počas 2 hodín pri teplote miestnosti, potom nasledovala inkubácia s myšacím sérom v zriedení 1:100 v PBS pri 4 °C cez noc. Nasledujúci deň sa membrány trikrát premyli v PBS počas 10 minút za jemného kývania. Potom sa membrány inkubovali s HRP-konjugovanými protilátkami (Abs, kozí anti-myší IgG) (Sigma-Aldrich, St. Louis, MO, USA) pri riedení 1:2000 v PBS počas 1 hodiny pri teplote miestnosti s jemným kývaním. Membrány sa trikrát premyli a detekcia protilátok sa uskutočnila chemiluminiscenciou s použitím substrátu pre Western blotting Pierce ECL (Bio-Rad, Hercules, CA, USA). Membrány sa inkubovali s činidlom ECL počas 3 minút a snímky membrán sa urobili pomocou systému ChemiDoc™ Touch Imaging System (Bio-Rad, Hercules, CA, USA).
Nepriama ELISA
Hladiny Abs reaktívnych proti bakteriálnym proteínom boli merané v myších sérach, ako už bolo uvedené [ 13 ]. 96-jamkové doštičky ELISA (Thermo Scientific, Waltham, MA, USA) boli potiahnuté 100 ul na jamku 0,5 ug/ml proteínových extraktov E. coli BL21 a inkubované počas 2 hodín pri teplote miestnosti s jemným kontinuálnym trepaním pri 100 ot./min. Následne boli doštičky inkubované cez noc pri 4 °C. Antigény sa zriedili v uhličitanovom/hydrogenuhličitanovom pufri (0,05 M, pH 9,6). Nasledujúci deň sa jamky trikrát premyli 100 ul PBS s obsahom 0,05 % (obj./obj.) Tween 20 (PBST) a potom sa blokovali pridaním 100 ul 1 % ľudského sérového albumínu (HSA)/PBS počas 1 hodiny pri teplote miestnosti a jemným kontinuálnym trepaním pri 100 ot./min. Po troch premytiach sa do jamiek pridali vzorky séra zriedené 1:700 v 0,5% HSA/PBS a inkubovali sa 1 hodinu pri 37 °C pri 100 ot./min. Doštičky sa trikrát premyli a pridalo sa 100 ul na jamku protilátok konjugovaných s HRP (kozí anti-myší IgG a IgM) (Sigma-Aldrich, St. Louis, MO, USA) v zriedení 1:1500 v 0,5% HSA/PBST a inkubovalo sa 1 hodinu pri teplote miestnosti pri 10 ot./min. Doštičky sa trikrát premyli a reakcia sa vyvolala so 100 ul roztoku TMB pripraveného na použitie (Promega, Madison, WI, USA) pri teplote miestnosti počas 20 minút v tme a potom sa zastavila s 50 ul 0,5 MH2S04 . Optimálna koncentrácia antigénu a riedenia séra a konjugátu boli definované pomocou titračného testu. Optická hustota (OD) sa merala pri 450 nm pomocou čítačky platní ELISA (Filter-Max F5, Molecular Devices, San Jose, CA, USA). Všetky vzorky boli testované trojmo a priemerná hodnota troch slepých pokusov (žiadne Abs) bola odčítaná od nameraných hodnôt. Hladiny Abs v predadsorbovaných sérach sa merali potiahnutím 100 ul na jamku 0,5 ug/ml proteínových extraktov B. afzelii a podľa vyššie opísaného protokolu.
Kliešťové kapilárne kŕmenie
Kapilárne kŕmenie sa uskutočňovalo pomocou nekŕmených nymf I. ricinus . Sklenené kapiláry s priemerom 3,5″ (Drummond Scientific, Broomall, PA, USA) sa naplnili roztokom obsahujúcim 5 x 106 spirochét B. afzelii v BSK-II samostatne alebo v kombinácii s 2,5 x 107 buniek /ml E. coli BL21. Podiel E. coli BL21 k B. afzelii (5:1) bol vybraný na základe množstva zisteného pre obe baktérie v neinfikovanej skupine. Kliešte sa pripevnili na plastové Petriho misky obojstrannou lepiacou páskou a naplnené kapiláry sa umiestnili na ústie kliešťa. Kliešte sa nechali kŕmiť 4 hodiny vo vlhkej komore pri 33 °C. Po kŕmení boli kliešte oddelené od obojstrannej pásky, zozbierané do sterilnej skúmavky s otvormi a udržiavané v inkubátore s > 97 % relatívnou vlhkosťou pri izbovej teplote počas 6 hodín pred extrakciou DNA.
Mikroinjekcia kliešťa
Mikroinjekčný experiment sa uskutočnil na nekŕmených nymfách. Mikroinjekčné kapiláry 3,5″ (Drummond Scientific, Broomall, PA, USA) boli vyrobené zahrievaním a ťahaním 1 mm sklenených kapilárnych trubíc v zariadení na vyťahovanie sklenených mikropipet (P-1000 Sutter Instrument, Novato, CA, USA). Pre nový experiment s pridaním komenzálnych baktérií sa na mikroinjekciu použila kultúra B. afzelii pestovaná v médiu BSK-II (5 x 106 spirochét /ml) samostatne alebo v kombinácii s E. coli BL21 (2,5 x 107 buniek/ml). Na adsorpčný experiment sa séra myší imunizovaných E. coli a myší infikovaných B. afzelii preinkubovali cez noc pri 4 °C s 250 ng/ul proteínov B. afzelii . Nasledujúci deň boli vopred adsorbované séra centrifugované pri plnej rýchlosti počas 30 minút a supernatant bol izolovaný. Potom sa na mikroinjekciu použila kultúra B. afzelii pestovaná v médiu BSK-II (5 x 106 spirochét/ml) samostatne alebo v kombinácii s PBS + B. afzelii , E. coli + B. afzelii alebo vopred adsorbovaným myšacím sérom E. coli + B. afzelii . Kliešte boli dočasne imobilizované na obojstrannej páske a mikroinjekcie boli uskutočnené pomocou mikrostriekačkovej pumpy (Drummond) pripojenej k Micro4 Controller (World Precision Instruments). Objem 8 nl bol vstreknutý do análneho póru nekŕmených nýmf. Mikroinjikované kliešte boli pred extrakciou DNA inkubované pri teplote miestnosti počas 2 alebo 6 hodín v inkubátore pri > 97 % relatívnej vlhkosti.
Extrakcia DNA
Genomická DNA bola extrahovaná z plne prekrvených lariev, nýmf a myších tkanív. DNA z jednotlivých plne prekrvených lariev bola extrahovaná 15 dní po kŕmení. DNA z jednotlivých nýmf bola extrahovaná na konci inkubačnej doby z kapilárneho kŕmenia a mikroinjekčných experimentov. DNA z myších tkanív sa extrahovala na konci experimentu (52. deň). Jednotlivé kliešte boli rozdrvené jednorazovou sondou, zatiaľ čo tkanivá myší boli rozdrvené sklenenými guľôčkami pomocou homogenizátora Precellys24 Dual (Bertin Technologies, Paríž, Francúzsko) pri 5500 x g počas 20 s dvakrát. Genómová DNA bola extrahovaná z tkanív kliešťov a myší pomocou súpravy na extrakciu DNA tkaniva Nucleospin (Macherey–Nagel, Hoerdt, Francúzsko). Každá vzorka DNA z kliešťov a myšacieho tkaniva sa eluovala v 20 a 50 ul sterilnej vody, v danom poradí. Kvalita genómovej DNA (OD260/280 medzi 1,8 a 2,0) sa merala pomocou NanoDrop™ One (Thermo Scientific, Waltham, MA, USA).
Extrakcia RNA a syntéza cDNA
Celková RNA sa extrahovala z rôznych tkanív myší pomocou činidla Trizol (DE3, Invitrogen, Carlsbad, CA, USA) podľa odporúčaní výrobcu. Získaná RNA bola reverzne transkribovaná na cDNA pomocou Superscript III (DE3, Invitrogen, Carlsbad, CA, USA) a bola použitá na detekciu patogénu pomocou RT-qPCR.
Detekcia a kvantifikácia záťaže B. afzelii pomocou PCR a qPCR
Na detekciu B. afzelii v celých larvách alebo v tkanivách myší sa uskutočnil predamplifikačný krok na zlepšenie detekcie patogénnej DNA alebo cDNA. Na tento účel bola celková DNA alebo cDNA vopred amplifikovaná pomocou PreAmp Master Mix (Fluidigm, CA, USA) podľa pokynov výrobcu. Primery zacielené na gén 23S rRNA pre Borrelia spp. (23S rRNA-F ‚GAGTCTTAAAAGGGCGATTTAGT‘, 23S rRNA-R ‚CTTCAGCCTGGCCATAAATAG‘) boli spojené spojením rovnakého objemu každého priméru na konečnú koncentráciu 200 nM. Reakcia sa uskutočnila v konečnom objeme 5 μl obsahujúcom 1 μl Perfecta Preamp 5X, 1,25 μl spojenej zmesi primérov, 1,5 μl destilovanej vody a 1,25 μl DNA. Program termocyklovania pozostával z jedného cyklu pri 95 °C počas 2 minút, 14 cyklov pri 95 °C počas 15 s a 4 minút pri 60 °C. Na konci cyklovacieho programu boli reakcie zriedené 1:2 v ultračistej vode Milli-Q. Následne sa uskutočnila kvantitatívna PCR (qPCR) s použitím rovnakých vyššie uvedených primérov a ďalšej sondy (23S rRNA-sonda ‚AGATGTGGTAGACCCGAAGCCGAGT‘) v LightCycler 480 (Roche, Meylan, Francúzsko). Reakčná zmes obsahovala 6 μl Master univerzálnej sondy FastStart (Roche), 0,12 μL 20 μM primérov 23S rRNA-F, 23S rRNA-R a TaqMan sonda 23S rRNA-sonda, 2 μl predamplifikovanej DNA alebo cDNA vzorky ultračistej vody až 2 Milli. Program amplifikácie pozostával z nasledovného: 95 °C počas 5 minút, 45 cyklov pri 95 °C počas 10 s a 60 °C počas 15 minút. Záťaž spirochét v kliešťoch bola získaná interpoláciou hodnoty CT v štandardnej krivke „počet spirochét vs CT“ a potom bola normalizovaná množstvom DNA v každej vzorke.
Detekcia Enterobacteriaceae pomocou PCR
DNA extrahovaná z celého nymfálneho kliešťa sa použila na detekciu Enterobacteriaceae pomocou nasledujúceho páru primérov: F-Enterobacteriaceae „ATGGCTGTCGTCAGCTCGT“, R-Enterobacteriaceae „CCTACTTCTTTTGCAACCCACTC“ (z [ 18 ]), ktoré sa zameriavajú na gén 16S rRNA pre Enter. Reakcia sa uskutočnila v konečnom objeme 50 μl obsahujúcom 5 μl 10X pufra, 4 μl dNTP, 1 μl každého primeru, 0,25 μl Taq polymerázy (Takara, Shiga, Japonsko), 1 μl DNA a 37,75 μl destilovanej vody. Zmesi sa amplifikovali počas 40 cyklov pri 98 °C počas 10 s, 55 °C počas 30 s a 72 °C počas 1 minúty, s konečným predĺžením pri 72 °C počas 3 minút v automatizovanom tepelnom cykléri (Perkin-Elmer Cetus, Gouda, Holandsko). Alikvóty obsahujúce 3 μl každého amplifikovaného produktu, 1 μl gélového nanášacieho pufra (Thermo Scientific, Waltham, MA, USA) a 2 μl destilovanej vody boli podrobené elektroforéze v 1,0 % (hmotn./obj.) agarózovom géli s paralelným markerom veľkosti molekuly (Thermo Scientific, Waltham, MA, USA). DNA z kultúry E. coli BL21 sa použila ako pozitívna kontrola. Elektroforéza v TAE (40 mM Tris-acetát, 1 mM EDTA) tlmivom roztoku (Lonza Biosciences, Basel, Švajčiarsko) sa uskutočnila pri 90 V počas 1,5 hodiny. Gél bol zafarbený pomocou GelGreen (Biotium, Fremont, CA, USA) a fotografovaný pri osvetlení ultrafialovým svetlom.
Príprava knižnice Illumina a sekvenovanie génu 16S rRNA
Najmenej 400 ng úplne prekrvenej DNA lariev v koncentrácii ≥ 20 ng/μl sa odoslalo na sekvenovanie amplikónov bakteriálneho génu 16S rRNA, ktoré bolo zadané spoločnosti Novogene Bioinformatics Technology Co. (Londýn, Spojené kráľovstvo). Knižnice boli pripravené pomocou súpravy NEBNext® Ultra™ IIDNA Library Prep Kit (New England Biolabs, MA, USA). Jedna dráha systému Illumina MiSeq sa použila na generovanie 251-bázových párových koncov čítaní z V4 variabilnej oblasti 16S rRNA génu pomocou čiarových kódovaných univerzálnych primerov (515F/806R) vo vzorkách z lariev nahromadených neinfikovanými myšami ( n = 10 ) , larva B. in engorged na E. coli imunizované a B. afzelii – infikované ( n = 8) alebo falošne imunizované a B. afzelii – infikované myši ( n = 10). Surové génové sekvencie 16S rRNA získané zo vzoriek kliešťov boli uložené v úložisku SRA (Bioproject No. PRJNA870490).
Kontroly, identifikácia a odstraňovanie kontaminantov
Boli nastavené dve kontroly extrakčného činidla, v ktorých sa rôzne kroky extrakcie DNA uskutočňovali s použitím rovnakých podmienok ako pre vzorky, ale s použitím vody ako templátu. Amplifikácia DNA sa potom uskutočnila na extrakčnej kontrole za rovnakých podmienok ako pri akejkoľvek inej vzorke. Možná kontaminujúca DNA vo vzorkách na sekvenovanie génu 16S rRNA bola štatisticky identifikovaná pomocou balíka „decontam“ [ 19 ] pomocou metódy „prevalencie“. Prevalencia je definovaná ako prítomnosť alebo neprítomnosť vo vzorke a použitá metóda porovnáva prevalenciu každého sekvenčného znaku v skutočných vzorkách s prevalenciou v negatívnych kontrolách na identifikáciu kontaminantov. Potom boli kontaminanty odstránené zo súboru údajov pred následnou analýzou mikrobiómov [ 19 ].
Analýza 16S rRNA génových amplikónových sekvencií
Analýza sekvencií génu 16S rRNA sa uskutočnila pomocou potrubia QIIME 2 (v. 2021.4) [ 20 ]. Pomocou softvéru DADA2 [ 21 ] implementovaného v QIIME2 boli génové sekvencie 16S rRNA najprv demultiplexované a potom kvalitatívne orezané na základe priemernej kvality na bázu pri doprednom a spätnom čítaní. Celková dĺžka bola skrátená o 180 a 154 pri čítaní dopredu a dozadu. Následne boli čítania zlúčené a chimérické varianty boli odstránené. Výsledné reprezentatívne sekvencie boli taxonomicky priradené pomocou vopred trénovaného naivného Bayesovho taxonomického klasifikátora [ 22 ] na základe verzie databázy SILVA 132 [ 23 ] a sady primérov 515F/806R. Výsledné tabuľky taxonomických údajov sa zbalili na úrovni rodu a taxóny s menej ako 10 celkovými čítaniami a prítomnými v menej ako 30 % vzoriek každého súboru údajov sa odstránili. Tabuľky taxonomických údajov sa použili na sieťovú analýzu a identifikáciu základných taxónov. Pre pohodlie v tejto štúdii odkazujeme na rod Borrelia ako na jeden rod v zmysle prezentovanom [ 24 ] a neuvažovali sme o rozdelení rodu Borrelia na dva rody: pozmenený rod Borrelia obsahujúci len členov recidivujúcej horúčky Borrelia a rod Borreliella obsahujúci členov komplexu lymskej choroby Borrelia] .25 dorferi (tj B.2 ) . To neznamená, že zaujímame akékoľvek stanovisko k súčasnej diskusii o tejto otázke [ 26 ].
Konštrukcia sietí spoločného výskytu baktérií, identifikácia kľúčových taxónov a test tolerancie útoku
Sieťové analýzy spoločného výskytu sa uskutočnili pomocou metódy Sparse Correlations for Compositional data (SparCC) [ 27 ] implementovanej v R studio [ 28 ]. Na výpočet korelačnej matice sa použili tabuľky taxonomických údajov. Boli zvolené korelačné koeficienty s veľkosťou > 0,75 alebo < − 0,75. Vizualizácia siete a výpočet topologických prvkov a prepojenosti taxónov (tj počet uzlov a hrán, modularita, priemer siete, priemerný stupeň, vážený stupeň, koeficient zhlukovania a metriky centrality) boli vykonané pomocou softvéru Gephi 0.9.2 [ 29 ]. Robustnosť sietí spoločného výskytu bola testovaná testom tolerancie útoku pomocou balíka NetSwan for R [ 30 ]. Na tento účel boli siete podrobené systematickému odstraňovaniu uzlov pomocou riadeného útoku, kde sú uzly odstraňované v klesajúcom poradí podľa hodnoty ich centrality medziľahlosti (BNC) (tj koľkokrát sa uzol nachádza na najkratšej ceste medzi ostatnými uzlami).
Porovnávacia sieťová analýza
Porovnanie podobnosti najcentrálnejších uzlov medzi dvoma sieťami sa uskutočnilo s balíkom „NetCoMi“ [ 31 ] v R studio s použitím taxonomických tabuliek počtu čítaní. „Najcentrálnejšie“ uzly sú definované ako uzly s hodnotou centrality nad empirickým 75 % kvartilom. Porovnanie vráti Jaccardove indexy pre každú zo štyroch lokálnych mier (tj stupeň, centrálnosť medziľahlosti, centrálnosť blízkosti, centrálnosť vlastného vektora) množín väčšiny centrálnych uzlov, ako aj množín centrálnych taxónov medzi dvoma porovnávanými sieťami. Jaccardov index teda vyjadruje podobnosť súborov väčšiny centrálnych uzlov, ako aj súborov hubových taxónov medzi týmito dvoma sieťami. Jaccardov index 0 označuje úplne odlišné súbory, zatiaľ čo hodnota 1 označuje rovnaké súbory väčšiny centrálnych uzlov alebo taxónov uzlov medzi porovnávanými sieťami [ 31 ].
Štatistická analýza
Tabuľka taxonomických údajov, ktorá pozostávala zo sekvenčných čítaní, sa použila ako vstup balíka R „ALDEx2“ [ 32 ], ktorý vykonal transformáciu centrovaného log-ratio (clr) pre všetky funkcie vo všetkých vzorkách. Početnosť taxónov sa porovnávala pomocou balíka R „DeSeq2“ [ 33 ]. Počet zdieľaných priamych susedov referenčného taxónu Escherichia-Shigella v rôznych experimentálnych skupinách bol vizualizovaný pomocou Vennových diagramov implementovaných v online nástroji http://bioinformatics.psb.ugent.be/webtools/Venn/ . Alfa a beta diverzita bakteriálnych taxónov sa uskutočnila na zriedkavých ASV tabuľkách. Alfa diverzita bola skúmaná pomocou Pielouovej rovnomernosti a fylogenetických metrík viery. Rozdiely v metrikách alfa diverzity medzi skupinami boli testované pomocou Kruskal-Wallisovho testu. Beta diverzita sa skúmala pomocou Jaccardovej podobnosti a vážených meraní Unifrac a porovnávala sa medzi skupinami pomocou testu PERMANOVA. Funkcia betadisper v R sa použila na určenie disperzie vzoriek na základe Bray-Curtisovej matice vzdialenosti a test analýzy rozptylu (ANOVA) sa použil na porovnanie disperzie vzoriek medzi skupinami. Na testovanie podobnosti väčšiny centrálnych uzlov sa vypočítali dve hodnoty p P(J ≤ j) a P(J ≥ j) pre každý Jaccardov index, ktoré predstavujú pravdepodobnosť, že pozorovaná hodnota Jaccardovho indexu je „menšia alebo rovná“ alebo „vyššia alebo rovná“ náhodne očakávanej Jaccardovej hodnote. Rozdiely v relatívnych hladinách Ab (tj OD) medzi skupinami imunizovaných myší v rôznych časových bodoch sa porovnávali s použitím dvojcestnej ANOVA s Bonferroniho viacnásobnými porovnávacími testami aplikovanými na individuálne porovnania. Rozdiely v relatívnych hladinách Ab po predadsorpcii s proteínmi B. afzelii sa porovnávali s použitím jednosmernej ANOVA s Tukeyho testom viacnásobného porovnávania. Rozdiely v záťaži patogénmi v skupinách infikovaných kliešťov, ktoré dostali vopred adsorbované séra v porovnaní s kontrolnými skupinami, boli analyzované pomocou Kruskal-Wallisovho testu. Zhluková analýza rôznych vzoriek bola založená na matici vzdialenosti Jaccard a bola vykonaná pomocou balíka „Vegan“ [ 34 ] v R pomocou Wardovej metódy. Nepárový neparametrický Mann–Whitney U test sa použil na porovnanie parametrov kliešťov (tj percento kliešťov, ktoré prirodzene klesli, percento lariev, ktoré sa zliali na nymfy a úmrtnosť kliešťov) a záťaže B. afzelii v kliešťoch medzi skupinami. Kruskal-Wallisov test s Dunnovým viacnásobným porovnávacím testom sa použil na porovnanie straty konektivity pri odstránení 5 až 7 % uzlov spomedzi všetkých experimentálnych podmienok. The Mann-Whitney Utest a Kruskal-Wallisov test nasledovaný Dunnovým viacnásobným porovnávacím testom sa uskutočnili v softvéri GraphPad 8 Prism (GraphPad Software Inc., San Diego, CA, USA). Rozdiely sa považovali za významné, keď p < 0,05.
Výsledky
B. afzelii moduluje mikrobiotu kliešťov
Na štúdium vplyvu infekcie B. afzelii na mikrobiotu kliešťov boli larvy I. ricinus kŕmené myšami infikovanými boréliami a neinfikovanými myšami (obr. 1 a). Následne sa analyzovalo zloženie bakteriálnej komunity a diverzita kliešťovej mikrobioty pomocou profilovania génu 16S rRNA po štatistickej identifikácii a odstránení znakov DNA (doplnková tabuľka S1 ) . Analýza indexov alfa diverzity ukázala, že fylogenetická diverzita Faith (obr. 2 a), ako aj rovnomernosť (obr. 2 b) sa nelíšili medzi kliešťami kŕmenými myšami infikovanými B. afzelii a neinfikovanými kliešťami (Kruskal–Wallis, p > 0,05). Analýza beta diverzity kliešťovej mikroflóry však odhalila, že infekcia B. afzelii viedla k posunu v zložení a početnosti bakteriálnej komunity v porovnaní s neinfikovanou skupinou, ako bolo merané pomocou Jaccardovho indexu (PERMANOVA, F = 1,84, p = 0,001, obr. 2 c) a váženej vzdialenosti unifrac (PERMAN, 34, F = 02 ) . resp. Okrem toho permutačný test na vyhodnotenie homogenity disperzií založený na matici vzdialenosti Bray-Curtis neodhalil žiadne významné rozdiely v disperzii bakteriálnej komunity medzi týmito dvoma skupinami ( F = 2,43, p > 0,05).
Obr.

Vplyv infekcie B. afzelii na zhromaždenie bakteriálnej komunity bol hodnotený vytvorením sietí mikrobiálneho spoločného výskytu. Vizuálna kontrola sietí ukázala, že infekcia B. afzelii spôsobila posun v zostavovaní bakteriálnych spoločenstiev (obr. 2e , f). Analýza topologických vlastností sietí odhalila zvýšený počet uzlov a najmä hrán v sieťach mikrobiálneho spoločného výskytu odvodených z mikrobioty kliešťov kŕmených na myšiach infikovaných B. afzelii v porovnaní so sieťou neinfikovanej skupiny (tabuľka 1 ). Podobne sa modularita a priemerný stupeň zvýšili v sieti B. afzelii v porovnaní s neinfikovanou sieťou (tabuľka 1 ). Pozorovaný Jaccardov index pre všetky testované lokálne miery centrality (tj stupeň, centralita medziľahlosti, centralita blízkosti, centralita vlastného vektora a taxóny uzla), s výnimkou centrality medziľahlosti, bol vyšší, ako sa očakávalo náhodne pre porovnania medzi neinfikovanými sieťami a sieťami B. afzelii (doplnková tabuľka S 2 ), čo naznačuje vysokú podobnosť uzlov v dvoch hierarchických organizáciách
Tabuľka 1.
Topologické vlastnosti sietí mikrobiálneho spoločného výskytu
| Topologické vlastnosti | Experimentálne skupiny | |
|---|---|---|
| Neinfikovaný | B. afzelii | |
| Uzly a | 626 (208) i | 645 (230) i |
| Okraje b | 474 | 1008 |
| -Pozitívne | 386 | 688 |
| -Negatíva | 88 | 320 |
| Modularita c | 0,908 | 1,476 |
| Moduly d | 56 | 45 |
| Priemer siete e | 11 | 11 |
| Priemerný stupeň f | 1,514 | 3.126 |
| Vážený stupeň g | 0,771 | 0,977 |
| Zhlukovací koeficient h | 0,409 | 0,568 |
a Uzly predstavujú bakteriálne taxóny s koreláciou spoločného výskytu SparCC > alebo < − 0,75
b hrany predstavujú počet spojení/korelácií
c modularita je sila rozdelenia siete na moduly
d moduly sú podspoločenstvá baktérií, ktoré sa medzi sebou vyskytujú častejšie ako s inými taxónmi
Priemer siete je najkratšia cesta medzi dvoma najviac oddelenými uzlami
f priemerný stupeň je priemerný počet odkazov na uzol
g vážený stupeň je súčet hmotnosti všetkých hrán pripojených k uzlu
h koeficient zhlukovania je miera, do akej uzly v sieti majú tendenciu vytvárať zhluky
i celkové uzly a uzly s aspoň jednou hranou sú v zátvorkách
Medzi kliešťami kŕmenými myšami infikovanými B. afzelii a neinfikovanou skupinou sa zistili významné zmeny v početnosti 65 taxónov (Wald test, p < 0,05, doplnkový obrázok S 1 ). Taxón Borrelia sa výrazne zmenil a jeho početnosť bola vyššia v mikrobiote kliešťov kŕmených myšami infikovanými B. afzelii (Wald test, p = 0,02, doplnkový obrázok S 1 ) v porovnaní s neinfikovanou skupinou. Spomedzi taxónov, ktorých početnosť sa výrazne zmenila, početnosť 51 a 14 bakteriálnych taxónov vzrástla, respektíve klesla u kliešťov zo skupiny B. afzelii . 14 taxónov s výrazne zníženou abundanciou v skupine B. afzelii je znázornených na obr. 2 g,h.
Je zaujímavé, že početnosť kľúčového taxónu Escherichia-Shigella sa významne znížila v skupine B. afzelii v porovnaní s neinfikovanou skupinou (Wald test, p = 0,03, doplnkový obrázok S 1 ). Rod Escherichia-Shigella bol opísaný ako kľúčový taxón v mikrobióme kliešťov Ixodes [ 13 , 14 ]. Okrem toho vakcínou indukované protilátky špecifické pre Escherichia-Shigella modulovali mikrobiotu kliešťa a vyvolali zmeny vo fyziológii kliešťa [ 13 ] a znížili vektorovú kompetenciu komárov pre P. relictum [ 15 ]. Vizuálna kontrola podsietí lokálnej interakcie Escherichia-Shigella ukázala, že infekcia B. afzelii zvýšila počet priamych susedov vyskytujúcich sa súčasne s Escherichia-Shigella v porovnaní s neinfikovanou podsieťou (obr. 2 i,j). Väčšina uzlov spojených s taxónom Escherichia-Shigella v neinfikovanej podsieti bola prítomná aj v podsieti B. afzelii (obr. 2 k, doplnková tabuľka S 3 ) a typ spojenia medzi nimi (tj pozitívna alebo negatívna korelácia) bol zachovaný (obr. 2 l). Našli sme tiež 23 jedinečných uzlov v podsieti B. afzelii , kde 12 a 11 uzlov má negatívnu a pozitívnu koreláciu spoločného výskytu s Escherichia-Shigella . Ďalšia charakterizácia dôležitosti rodu Escherichia-Shigella v sieťach spoločného výskytu odhalila zvýšenú centralitu blízkosti, centralitu medzisústrednosti a centrálnosť vlastného vektora (tabuľka 2 ). Pozoruhodné je, že mierou, ktorá sa najviac zmenila, bola centrálnosť medzi kliešťami, kde sa zvýšila šesťkrát v sieti kliešťov kŕmených myšami infikovanými B. afzelii v porovnaní s kliešťami kŕmenými neinfikovanými myšami. Tieto výsledky ukazujú, že napriek tomu, že početnosť Escherichia-Shigella je nižšia u kliešťov infikovaných B. afzelii , pravdepodobne v dôsledku zvýšenia bakteriálnej konkurencie, význam tohto taxónu v sieťach rastie. Celkovo výsledky ukázali, že infekcia B. afzelii viedla k posunu v mikrobiote kliešťov charakterizovanom zmenami v diverzite beta, bakteriálnou abundanciou, niektorými sieťovými vlastnosťami a relatívnou dôležitosťou Escherichia-Shigella .
Tabuľka 2
Miery centrálnosti taxónu Escherichia-Shigella v neinfikovaných sieťach a sieťach B. afzelii
| Experimentálne skupiny | Centrálnosť blízkosti a | Medzicentrálnosť b | Centrálnosť vlastného vektora c |
|---|---|---|---|
| Neinfikovaný | 0,256 | 0,001 | 0,635 |
| B. afzelii | 0,350 | 0,006 | 0,903 |
Centrálnosť blízkosti udáva, ako blízko je uzol ku všetkým ostatným uzlom v sieti
b Centrálnosť medzi uzlami udáva, nakoľko je daný uzol medzi ostatnými
Centrálnosť vlastného vektora meria dôležitosť uzla, pričom zohľadňuje dôležitosť jeho susedov
Antimikrobiota vakcína mení posun mikroflóry kliešťov vyvolaný B. afzelii a znižuje infekciu patogénom u kliešťov
Vyššie uvedené výsledky a predchádzajúce dôkazy [ 9 ] nás viedli k hypotéze, že zmena stavov permisívnych voči boréliám mikrobioty kliešťov by mohla zmeniť kolonizáciu patogénom vo vektore kliešťov. Aby sme otestovali túto hypotézu, zmenili sme mikrobiotu kliešťov zacielením na kľúčový taxón Escherichia-Shigella [ 14 ] v kliešťoch infikovaných B. afzelii a merali sme vplyv na zdatnosť patogénu. Po imunizácii živou vakcínou na báze E. coli nasledovala experimentálna infekcia B. afzelii a následné napadnutie kliešťami na myšiach (obr. 1 b). Infekcia B. afzelii bola potvrdená pomocou qPCR (doplnková tabuľka S 4 a, b) a RT-qPCR (doplnková tabuľka S 4 c, d) v rôznych tkanivách myší a pomocou western blotu s použitím myšacích sér proti proteínovým extraktom B. afzelii (doplnkový obrázok S 2 ). Vakcinácia myší s E. coli vyvolala imunitnú odpoveď, kde boli pozorované zvýšené hladiny protilátok IgM (obr. 3 a) a IgG (obr. 3 b) špecifických pre E. coli v sére myší v porovnaní s kontrolnou skupinou, ktorá dostala falošnú vakcínu. Táto imunitná odpoveď sa udržala v priebehu času, aspoň 52 dní po prvej imunizácii (obr. 3 ).
Obr.

Rozdiely v mikroflóre kliešťov kŕmených na myšiach infikovaných B. afzelii a imunizovaných antimikrobiotami alebo falošnou vakcínou boli hodnotené porovnaním alfa a beta diverzity bakteriálnych spoločenstiev. Vakcinácia E. coli nemala významný vplyv na bakteriálnu diverzitu (Kruskal–Wallis, p > 0,05, obr. 4 a) ani na druhovú vyrovnanosť (Kruskal–Wallis, p > 0,05, obr. 4 b). Podobne beta diverzita neodhalila samostatnú klastrizáciu experimentálnych skupín meranú pomocou Jaccardovho indexu (PERMANOVA, p > 0,05, obr. 4 c) a váženého unifracu (PERMANOVA, p > 0,05, obr. 4 d).
Obr.

Vizuálna kontrola sietí mikrobiálneho spoločného výskytu skonštruovaných z mikrobioty kliešťov kŕmených myšami PBS + B. afzelii a E. coli + B. afzelii ukázala, že vakcína proti mikrobiálnej flóre modulovala zostavenie bakteriálnej komunity (obr. 4 e, f), čo bolo ďalej potvrdené topologickými znakmi týchto sietí (tabuľka 3 ). Konkrétne sa počet kladných a záporných hrán výrazne zvýšil v sieťach E. coli + B. afzelii v porovnaní so sieťou PBS + B. afzelii (tabuľka 3 ). Podobne sa v sieti E. coli + B. afzelii v porovnaní s jej kontrolou zvýšili ďalšie topologické vlastnosti, ako je modularita, počet modulov, priemerný stupeň a priemer siete (tabuľka 3 ). Testovanie Jaccardovho indexu pre miery centrality lokálnej siete odhalilo, že stupeň (Jacc = 0,389, p = 0,03) a centralita blízkosti (Jacc = 0,404, p = 0,009) mali hodnoty Jacc vyššie, ako sa náhodne očakávalo pre porovnania medzi týmito dvoma sieťami (doplnková tabuľka S5 ) . Diferenciálna analýza početnosti každého taxónu v mikrobiote kliešťov ukázala, že početnosť 46 bakteriálnych taxónov sa významne zmenila medzi kliešťami skupín PBS + B. afzelii a E. coli + B. afzelii (Wald test, p < 0,05 , doplnkový obrázok S 3 ). Konkrétne sa početnosť 23 bakteriálnych taxónov zvýšila a tiež sa znížilo 23 taxónov v mikrobiote kliešťov kŕmených imunizovanými a B. afzelii -infikovanými myšami v porovnaní s kontrolnou skupinou. Top 20 taxónov s najvyššími rozdielmi násobných zmien medzi týmito dvoma skupinami je znázornených na obr. 4 g a sú uvedené na obr. 4 h.
Tabuľka 3.
Topologické vlastnosti sietí mikrobiálneho spoločného výskytu
| Topologické vlastnosti | Experimentálne skupiny | |
|---|---|---|
| PBS + B. afzelii | E. coli + B. afzelii | |
| Uzly | 739 (204) | 735 (378) |
| Hrany | 1124 | 2002 |
| -Pozitívne | 867 | 1421 |
| -Negatíva | 257 | 581 |
| Modularita | 0,963 | 1.1 |
| modul | 28 | 73 |
| Priemer siete | 11 | 12 |
| Priemerný stupeň | 3,042 | 5,448 |
| Vážený stupeň | 1,384 | 1,951 |
| Klastrovací koeficient | 0,590 | 0,471 |
Aby sa zistilo, či imunizácia živými baktériami mala vplyv na dôležitosť E. coli v mikrobióme kliešťov, vytvorili sa podsiete zložené z taxónu Escherichia-Shigella a priameho suseda. Porovnanie podsietí odhalilo, že počet súbežne sa vyskytujúcich taxónov s taxónom Escherichia-Shigella klesol v sieťach mikrobioty kliešťov kŕmených myšami imunizovanými E. coli a myšami infikovanými B. afzelii (obr. 4 j) v porovnaní s tými, ktoré sa kŕmili falošne imunizovanými a B. infectedce mizelii ( obr . Porovnanie taxonomickej identity priamych susedov medzi podsieťami PBS + B. afzelii a E. coli + B. afzelii ukázalo, že priamo sa vyskytujúce taxóny rodu Escherichia-Shigella boli väčšinou jedinečné pre každú experimentálnu podmienku a iba 12 bolo zdieľaných medzi podsieťami (obr. 4 k, príloha 6 ). Detailným porovnaním typu korelácie spoluvýskytu medzi taxónom Escherichia-Shigella a bežnými taxónmi medzi oboma podsieťami sa zistilo, že typ spojenia sa zachoval v podsieťach PBS + B. afzelii a E. coli + B. afzelii (obr. 4 l). Na rozdiel od zvýšenia miery centrálnosti taxónu Escherichia-Shigella v sieti B. afzelii v porovnaní s neinfikovanou skupinou sme pozorovali pokles vo všetkých troch mierach centrálnosti (tj blízkosť, medzisúrodnosť a centrality vlastného vektora) v mikroflóre kliešťov kŕmených E. coli- imunizovanými a B. afzelii infikovanými a B. afzelii. afzelii -infikované myši (tabuľka 4 ).
Tabuľka 4.
Miery centrality taxónu Escherichia-Shigella v skupinách PBS + B. afzelii a E. coli + B. afzelii
| Experimentálne skupiny | Centrálnosť blízkosti | Medzisústrednosť | Centrálnosť vlastného vektora |
|---|---|---|---|
| PBS + B. afzelii | 0,431 | 0,004 | 1 |
| E. coli + B. afzelii | 0,295 | 0,003 | 0,898 |
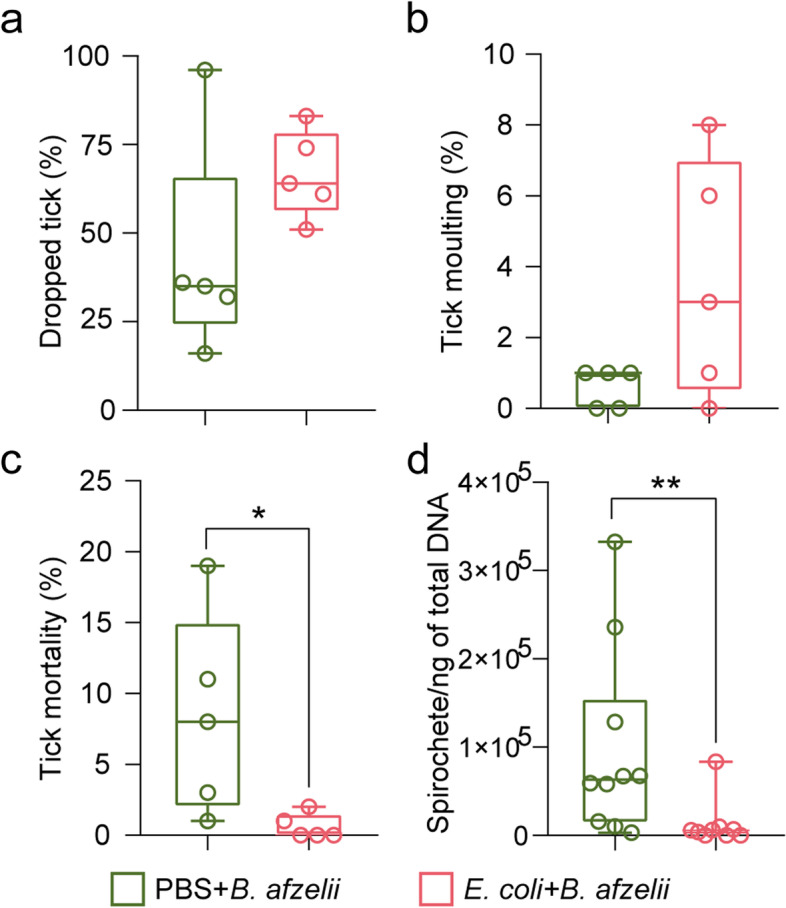
Na testovanie vplyvu antimikrobiotickej vakcíny na fyziológiu kliešťov a kondíciu patogénov sa porovnávalo niekoľko parametrov výkonnosti kliešťov a hladiny borélií medzi skupinami PBS + B. afzelii a E. coli + B. afzelii . Výsledky nepreukázali žiadne významné rozdiely v percentách kliešťov, ktoré prirodzene klesali (obr. 5 a), ani v percentách lariev, ktoré sa zliali na nymfy (obr. 5 b) medzi týmito dvoma skupinami. Bol však pozorovaný významný pokles úmrtnosti kliešťov zo skupiny E. coli + B. afzelii v porovnaní s kliešťami zo skupiny PBS + B. afzelii (Mann–Whitney test, p = 0,02, obr. 5 c). Je zaujímavé, že hladiny B. afzelii u kliešťov kŕmených E. coli -imunizovanými a B. afzelii -infikovanými myšami boli tiež významne nižšie ako u kliešťov kŕmených falošne imunizovanými a B. afzelii -infikovanými myšami (Mann–Whitney test, p = 0,0056, obr. 5 d). Tu sme ukázali, že antimikrobiálne vakcíny môžu narušiť mikrobiotu kliešťov a znížiť kolonizáciu boréliami , zatiaľ čo predchádzajúce štúdie ukázali, že narušenie mikrobioty kliešťov sterilným chovom kliešťov alebo vystavením antibiotikám tiež znížilo kolonizáciu boréliami [ 9 ].
Obr.
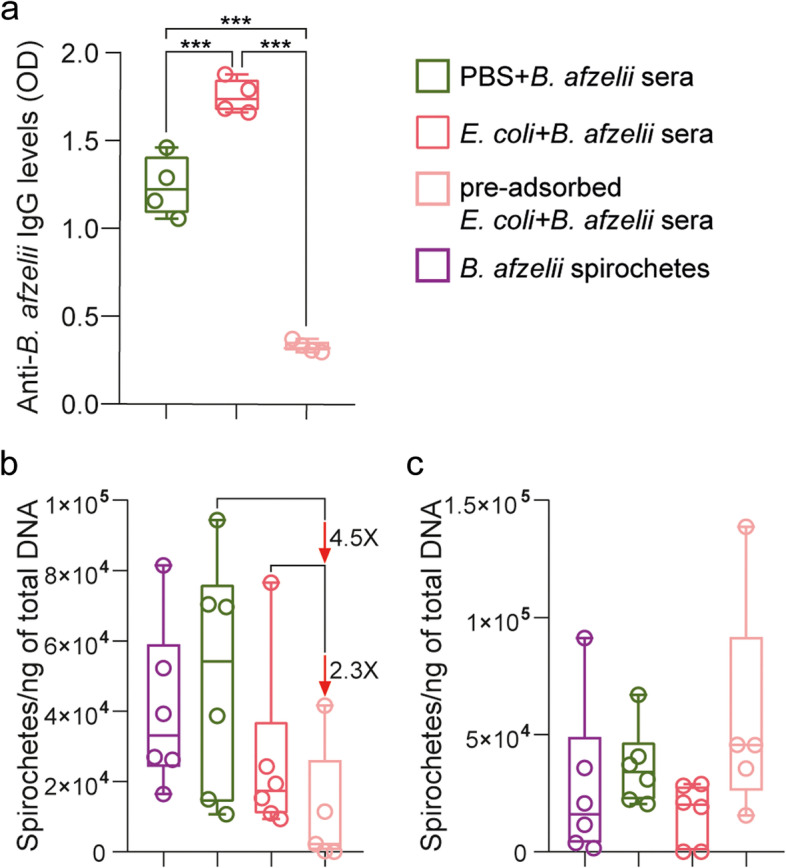
Predchádzajúce štúdie ukázali skríženú reaktivitu anti- E. coli protilátok proti boréliovým antigénom [ 35 , 36 ]. Aby sme vylúčili možnosť, že antimikrobiotové vakcíny znižujú záťaž boréliami iba v dôsledku efektu skríženej reaktivity anti- E. coli protilátok, odstránili sme protilátky anti- E. coli , ktoré by mohli skrížene reagovať s B. afzelii , a študovali sme vplyv zostávajúcich protilátok na záťaž patogénom v kliešťoch. Predpokladali sme, že zostávajúce protilátky anti- E. coli budú zamerané na baktérie Escherichia prítomné v mikrobiote kliešťov a ovplyvnia záťaž patogénom. Na to sme vopred adsorbovali séra myší imunizovaných E. coli a myší infikovaných B. afzelii antigénmi B. afzelii, aby sa odstránili krížovo reagujúce protilátky. Adsorpcia s proteínmi Borrelia významne znížila hladiny protilátok rozpoznávajúcich B. afzelii v sérach E. coli + B. afzelii v porovnaní s vopred neadsorbovanými sérami E. coli + B. afzelii alebo PBS + B. afzelii (ANOVA, p < 0,05, Obr. 6 a). Mikroinjekcia vopred adsorbovaných sér E. coli + B. afzelii do análneho póru infikovaných kliešťov znížila 4,5-násobok a 2,3-násobok záťaže B. afzelii v porovnaní s kliešťami mikroinjikovanými vopred neadsorbovaným PBS + B. afzelii alebo E. coli + B. afzelii séra > 0 , resp . obr . b) po 2 hodinách inkubácie. Pozorovali sme však, že tieto zníženia záťaže patogénmi v mikroinjikovaných kliešťoch boli prechodné a účinok vymizne po 6 hodinách inkubácie (obr. 6 c). Tieto výsledky naznačujú, že hoci prechodne, protilátky proti E. coli produkované vakcínou proti mikrobióze znížili záťaž boréliami nepriamo prostredníctvom modulácie mikrobioty kliešťov.
Obr.
Pridanie nových komenzálnych baktérií znižuje záťaž B. afzelii v nymfách I. ricinus
Aby sme otestovali, či iné spôsoby narušenia mikrobioty (tj pridanie novej komenzálnej baktérie, [ 37 – 42 ]) tiež znížili kolonizáciu boréliami , použili sme umelé kapilárne kŕmenie a mikroinjekciu do análneho póru na súčasné zavedenie E. coli BL21 a B. afzelii do nymfy I. ricinus . Použitím E. coli BL21, kmeňa dlho uchovávaného v laboratórnych podmienkach [ 43 , 44 ], sme zabezpečili novú interakciu v hostiteľovi kliešťa, čo nám umožnilo vylúčiť evolučnú históriu medzi pridanou baktériou, kliešťom a boréliou , čo mohlo znížiť jej vplyv na mikrobiotu.
Alfa a beta diverzita bakteriálnej komunity kliešťov, ktoré umelo dostali B. afzelii alebo E. coli + B. afzelii, sa analyzovala pomocou profilovania génu 16S rRNA po štatistickej identifikácii a odstránení znakov DNA (doplnková tabuľka S7 ) . Analýza mikrobioty ukázala, že súčasné pridanie E. coli a B. afzelii mikroinjekciou (ale nie s kapilárnym kŕmením) významne znížilo Faithovu fylogenetickú diverzitu a rovnomernosť (Kruskal–Wallis, p < 0,05, doplnkový obrázok S 4 a, b) mikrobiálnej komunity kliešťov v porovnaní s mikrobiálnym spoločenstvom kliešťov B. Okrem toho pridanie E. coli + B. afzelii mikroinjekciou alebo kapilárnym kŕmením významne zmenilo bakteriálne zloženie alebo početnosť mikrobioty kliešťov v porovnaní s kliešťami, ktoré dostali B. afzelii (PERMANOVA, p < 0, 05, doplnkový obrázok S 4 c, d).
Po 6 hodinách inkubácie sme porovnali záťaže patogénmi v kliešťoch vystavených E. coli a B. afzelii s tými, ktoré boli vystavené iba spirochétám B. afzelii . Prítomnosť pridanej E. coli v nymfách po kapilárnom kŕmení alebo mikroinjekcii bola potvrdená pomocou PCR (doplnkový obrázok S5 ) . Hladiny B. afzelii boli 2,4 (Mann–Whitney test, p ˃ 0,05, obr. 7 a) krát nižšie u nymf kŕmených z kapiláry E. coli v porovnaní s nymfami kŕmenými iba B. afzelii . Pre nymfy mikroinjikované E. coli a B. afzelii a inkubované 6 hodín sa záťaž B. afzelii znížila 2,0-krát v porovnaní s nymfami, ktorým bola injikovaná iba B. afzelii (Mann–Whitney test, p = 0,01, obr. 7 b).
Obr. 7.

To naznačuje, že Borrelia je vysoko citlivá na perturbácie kliešťovej mikrobioty a že odchýlky od modulácie vyvolanej patogénom vo vektorovej mikrobiote predstavujú pre spirochétu vysoké náklady.
Definovanie stavov odolných voči infekcii B. afzelii v mikrobiote I. ricinus
Uskutočnilo sa globálne porovnanie medzi štyrmi rôznymi experimentálnymi podmienkami (tj neinfikované, B. afzelii , PBS + B. afzelii a E. coli + B. afzelii) s cieľom definovať ekologické vlastnosti stavov odolných voči infekcii. Najprv sme porovnali taxonomický profil medzi všetkými experimentálnymi skupinami a zistili sme, že vysoký počet bakteriálnych taxónov (tj 535 taxónov) bol rozdelený medzi 4 skupiny. Iba 2 až 9 taxónov sa našlo výlučne v každom experimentálnom stave (obr. 8a ). Pri porovnaní zoznamu taxónov, ktorých početnosť sa významne zmenila medzi neinfikovanými skupinami a skupinami B. afzelii (doplnkový obrázok S 1 ) a medzi skupinami PBS + B. afzelii a E. coli + B. afzelii (doplnkový obrázok S 3 ), sme pozorovali, že iba 8 taxónov významne zmenilo svoju neinfikovanú abundanciu P. B. + porovnanie B. afzelii a E. coli + B. afzelii . Je zaujímavé, že väčšina taxónov bola jedinečná pre každý súbor porovnaní, kde 38 taxónov bolo nájdených výlučne v porovnaní medzi neinfikovanou skupinou a skupinou B. afzelii a 57 v porovnaní medzi skupinami PBS + B. afzelii a E. coli + B. afzelii (obr. 8 b). Okrem toho zhluková analýza mikrobioty založená na Jaccardovom indexe ukázala, že vzorky z rôznych experimentálnych podmienok sa zoskupili do troch rôznych skupín (obr. 8 c, doplnkový obrázok S 6 ). Každý zhluk tvorili väčšinou vzorky kliešťovej mikrobioty z jedného experimentálneho stavu, a to skupiny E. coli + B. afzelii , neinfikované a skupiny B. afzelii . Vzorky z PBS + B. afzelii sa zoskupili do dvoch rôznych skupín, so vzorkami zo skupín B. afzelii alebo E. coli + B. afzelii . Je pozoruhodné, že všetky vzorky kliešťovej mikroflóry zo skupiny E. coli + B. afzelii sa zoskupili do jednej oddelenej skupiny, čo neplatí pre ostatné experimentálne podmienky.
Obr. 8.

Následne sme porovnali vlastnosti vznikajúce v sieťach, aby sme určili ich možný príspevok k stavu odolnému voči infekcii B. afzelii v mikrobiote kliešťov. Porovnanie identity uzlov, ktoré sa podieľajú na interakciách mikróbov a mikróbov, ukázalo, že 85 bakteriálnych taxónov je spoločných v sieťach všetkých experimentálnych skupín (obr. 8 d). Je zaujímavé, že sieť, ktorá predstavovala najvyšší počet jedinečných taxónov (tj 157 uzlov), bola odvodená zo skupiny E. coli + B. afzelii v porovnaní s neinfikovanými (tj 22 uzlov), B. afzelii (tj 35 uzlov), PBS + B. afzelii (tj 13 uzlov). Navyše, aby sme určili, aká veľká časť bakteriálnej diverzity je preložená do interakcií mikróbov a mikróbov, porovnali sme pozorované znaky oproti počtu uzlov (obr. 8 e) alebo hrán (obr. 8 f), ktoré sa nachádzajú v sieťach mikrobiálneho spoločného výskytu. Pozorovali sme, že neinfikovaná skupina B. afzelii a PBS + B. afzelii s podobným počtom pozorovaných znakov predstavovala podobný počet interagujúcich uzlov v sieťach mikrobiálneho spoločného výskytu (obr. 8e ). Naopak, pre podobný počet pozorovaných znakov sa počet hrán zvýšil dvakrát v skupine B. afzelii a PBS + B. afzelii v porovnaní s neinfikovanou skupinou (obr. 8 f). Je zaujímavé, že skupina E. coli + B. afzelii s najvyššou bakteriálnou diverzitou mala aj najvyšší počet interagujúcich uzlov a bakteriálnych asociácií (obr. 8 f). Nakoniec sme testovali robustnosť sietí meraním ich tolerancie voči riadenému odstraňovaniu taxónov a porovnávali sme to medzi všetkými experimentálnymi skupinami. Sieť odvodená zo skupiny E. coli + B. afzelii vykazovala najnižšie hodnoty straty konektivity po priamom odstránení uzlov (obr. 8 g), čo naznačuje najvyššiu toleranciu na odstránenie taxónov v tejto sieti. Štatistické porovnanie straty konektivity pre 5 až 7 % odstránených uzlín ukázalo významný rozdiel medzi skupinami E. coli + B. afzelii a neinfikovanými skupinami B. afzelii alebo PBS + B. afzelii (obr. 8 g). Všetky tieto výsledky ukázali, že významné zmeny jedinečných bakteriálnych taxónov, vysoké mikrobiálno-mikrobiálne interakcie s unikátnym súborom uzlov, ako aj vyššia robustnosť mikrobiálnych sietí môžu definovať stavy odolné voči infekcii Borrelia v mikrobiote I. ricinuskŕmené imunizovanými myšami.
Pri porovnaní mikrobioty kliešťov, ktoré dostali umelo B. afzelii alebo E. coli + B. afzelii kapilárnym kŕmením alebo mikroinjekciou, sme zistili určité podobnosti s výsledkami uvedenými vyššie, pokiaľ ide o vysoký počet bakteriálnych taxónov (tj 470), ktoré zdieľajú všetky experimentálne skupiny (doplnkový obrázok S 7 a); vysoký počet jedinečných taxónov, ktorých početnosť sa výrazne zmenila v kapilárnych kŕmnych (tj 21) alebo mikroinjekčných (tj 31) skupinách medzi B. afzelii a E. coli + B. afzelii (doplnkový obrázok S 7 b), zhlukovanie väčšiny vzoriek z experimentálnych podmienok v oddelenom zhluku (doplnkový obrázok 7 c). Navyše, pri porovnaní uzlov prítomných v sieťach mikrobiálneho spoločného výskytu sme na rozdiel od vyššie uvedených výsledkov zistili, že skupina s najvyšším počtom jedinečných uzlov bola skupina, ktorá dostala B. afzelii kapilárnym kŕmením (doplnkový obrázok S 7 d). Keď sme porovnali pozorované znaky vs počet uzlov alebo okrajov, zistili sme, že stav E. coli + B. afzelii zo skupín s kapilárnym kŕmením alebo mikroinjekciou mal znížený počet uzlov (doplnkový obrázok S 7 e) a okrajov (doplnkový obrázok S 7 f) v porovnaní s ich kontrolnou skupinou ( stav B. afzelii ). Nakoniec sme pozorovali zníženie robustnosti mikrobiálnych sietí v podmienkach E. coli + B. afzelii zo skupiny s mikroinjekciou v porovnaní so skupinou B. afzelii . Neboli zistené žiadne významné rozdiely medzi podmienkami E. coli + B. afzelii a B. afzelii z mikroinjekčnej skupiny (doplnkový obrázok S 7 g).
Tieto výsledky ukazujú, že odchýlky od modulácie mikrobioty vyvolanej B. afzelii môžu ovplyvniť kolonizáciu patogénom v kliešťoch I. ricinus a že stav odolný voči infekcii závisí od techník používaných na narušenie mikrobioty.
Diskusia
Perturbácia mikroflóry je sľubnou cestou na vývoj opatrení na kontrolu chorôb prenášaných vektormi, pretože vektorová mikroflóra ovplyvňuje kompetenciu vektora [ 45 – 47 ]. Tu sme najprv charakterizovali interakciu kliešťovej mikroflóry s patogénom B. afzelii a následne sme ukázali, ako narušenie patogénom indukovanej mikroflóry viedlo k stavu odolnému voči infekcii, ktorý redukoval kolonizáciu boréliami vo vektoroch kliešťov.
Iné štúdie zistili, že prítomnosť borélií vo vektore výrazne zmenila mikrobiotu kliešťov. Napríklad nymfy I. ricinus vyliate z lariev, ktoré sa živili myšami infikovanými B. afzelii, predstavovali menej hojný, ale rozmanitejší bakteriálny mikrobióm [ 48 ]. Medzi kliešťami infikovanými a neinfikovanými B. afzelii boli tiež zistené rozdielne abundancie niekoľkých taxónov [ 48 ]. Na rozdiel od toho štúdia Chauhana a spol. [ 49 ] nezistili žiadnu súvislosť medzi diverzitou mikrobiómov kliešťa a jeho pravdepodobnosťou nosičstva B. burgdorferi , ale špecifické mikrobiálne taxóny boli spojené s prítomnosťou patogénu v jednotlivých kliešťoch. Tieto výsledky naznačujú, že infekcia Borrelia vyvolala modifikácie mikrobioty kliešťov spojené najmä so zmenami v množstve bakteriálnych členov. Naše výsledky tiež ukázali, že B. afzelii spôsobuje veľké posuny v zostave bakteriálnej komunity a zvyšuje spoločný výskyt baktérií, čo naznačuje vyššiu mieru interakcií mikróbov a mikróbov u infikovaných kliešťov. V minulosti bola tiež publikovaná modifikácia asociácií bakteriálnych taxónov vyvolaná prítomnosťou rôznych patogénov rodov Borrelia , Anaplasma a Rickettsia [ 50 ]. Na rozdiel od našich výsledkov však Lejal et al. [ 50 ] zistili, že bakteriálne korelácie boli nižšie v sieti z kliešťov pozitívnych na borélie v porovnaní so sieťou z neinfikovaných kliešťov. Tieto rozdiely by mohli byť spôsobené druhmi Borrelia , o ktorých sa uvažovalo. V štúdii Lejal et al. [ 50 ] sa vzorky považovali za pozitívne na borélie , ak boli detegované mikrofluidnou PCR a bakteriálne taxóny rovnakých rodov boli detegované sekvenovaním génu 16S rRNA. V ich štúdii je teda druh Borrelia neznámy. Rôzne druhy by mohli viesť k rôznym výsledkom, ale na potvrdenie tejto hypotézy sú potrebné ďalšie porovnávacie štúdie o vplyve rôznych druhov patogénov na mikrobiotu kliešťov. Napriek rozdielom medzi štúdiami dôkazy konzistentne ukazujú, že prítomnosť patogénu vo vektore moduluje vektorovú mikroflóru smerom k stavu kompatibilnému s kolonizáciou patogénom.
Vzhľadom na to, že patogény, ako sú borélie , musia prekonať niekoľko prekážok (napr. vyhnúť sa imunitnej obrane kliešťov alebo sa vyhnúť endocytickému tráveniu v epiteliálnych bunkách čreva kliešťa), aby pretrvali v kliešťoch až do ďalšej krvnej potravy, musia aktivovať mechanizmy na moduláciu prostredia čreva, aby sa uľahčila ich kolonizácia a pretrvávanie vo vektore [ 3 ]. Mechanicky sa ukázalo, že B. burgdorferi indukuje expresiu črevného génu I. scapularis kódujúceho PIXR (proteín I. scapularis s reelerovou doménou) [ 51 ]. Zrušenie funkcie PIXR in vivo viedlo k zmenám v črevnom mikrobióme, metabolóme a imunitných odpovediach a knockdown PIXR sprostredkovaný RNA interferenciou znížil kolonizáciu B. burgdorferi , čo naznačuje, že B. burgdorferi indukuje expresiu PIXR na zvýšenie kolonizácie v kliešťoch [ 51 ]. Podobne Anaplasma phagocytophilum , pôvodca ľudskej granulocytárnej anaplazmózy, indukuje expresiu génu kódujúceho nemrznúci glykoproteín Ixodes scapularis (IAFGP), ktorý narúša črevnú mikroflóru kliešťa a integritu peritrofickej matrice a črevnej bariéry s cieľom uľahčiť infekciu [ 52 ]. Široké účinky, ktoré má patogén na črevá kliešťov, môžu vytvoriť prostredie, ktoré uprednostňuje alebo znižuje kondíciu niektorých baktérií, čo môže vysvetliť zmeny v diverzite, zložení alebo množstve taxónov v mikrobiote. Okrem toho by modulácia mikrobioty mohla viesť k vymiznutiu alebo vzniku mikrobiálno-mikrobiálnych interakcií. V súlade s tým sme zistili, že kľúčový taxón Escherichia-Shigella [ 13 ] bol asociovaný s viacerými bakteriálnymi taxónmi v sieti B. afzelii v porovnaní s kontrolnou sieťou. Pochopenie presnej úlohy týchto novo vytvorených asociácií na perzistenciu patogénu vo vektore si však vyžaduje ďalšie skúmanie.
Vektorová mikrobiota sa považuje za bránu prístupu k vektorovej zdatnosti a kompetencii [ 53 ]. Vyvolanie zmien v normálnej mikrobiote kliešťov môže viesť k narušeniu kolonizácie patogénom [ 9 ]. Tu sme ukázali, že kliešte kŕmené na myšiach imunizovaných baktériami Enterobacteriaceae E. coli BL21 vykazovali nižšiu úroveň záťaže B. afzelii vo vektore. Tieto výsledky sú v súlade s predchádzajúcou štúdiou, kde komáre kŕmené imunizovanými vtákmi proti dvom kmeňom E. coli , konkrétne O86:B7 a BL21, mali znížený počet oocýst P. relictum (kauzálny agens vtáčej malárie) v stredných črevách [ 15 ]. Zníženie infekčnosti Plasmodium bolo spôsobené zmenou Plasmodium -indukovanej modulácie mikrobioty komárov [ 15 ]. Podobne sme v tejto štúdii zistili, že antimikrobiálna vakcína vyvolala zmeny v zostave mikrobiálnej komunity odlišné od tých, ktoré vyvolala B. afzelii . Zaujímavé je, že sme tiež zistili, že antimikrobiotová vakcína znížila relatívnu dôležitosť taxónu Escherichia-Shigella v porovnaní s rovnakým taxónom v skupine PBS + B. afzelii . Tieto výsledky naznačujú, že zacielenie na Enterobacteriaceae s hostiteľskými protilátkami indukuje moduláciu vektorovej mikroflóry, čo spúšťa kaskádový ekologický dopad na celú mikrobiotu kliešťa, čo viedlo k narušeniu kolonizácie patogénom. Podporujúc túto myšlienku, Narasimhan a kol. [ 9 ] uviedli, že experimentálne vyvolaná dysbiózna mikroflóra spôsobená chovom kliešťov v sterilnom prostredí znížila kolonizáciu B. burgdorferi ss v strednom čreve kliešťa. Tiež sa uvádza, že tento výsledok bol spojený s nižšou expresiou transkripčného faktora STAT a glykoproteínu peritrofínu, ktorý ovplyvnil integritu peritrofickej matrice, čo je nevyhnutné pre úspešnú kolonizáciu B. burgdorferi ss . Okrem toho Rana a spol. [ 54 ] preukázali, že IFNγ, prítomný u myší infikovaných boréliou , môže aktivovať dráhu Dome1-JAK-STAT, ktorá by mohla ovplyvniť mikrobiotu kliešťov, kolonizáciu patogénom a prenos na naivného hostiteľa. Súčasné získanie hostiteľských faktorov (napr. IFNγ) a antimikrobiálnych protilátok indukovaných imunizáciou v kontexte boréliovej infekcie by mohlo vysvetliť niektoré z rozdielov, ktoré sme pozorovali medzi mikroflórou kliešťov kŕmených infikovanými myšami v porovnaní s kliešťami kŕmenými kapilárou alebo mikroinjikovanými. Či sú tieto mechanizmy zapojené do účinkov antimikrobiotickej vakcíny, zostáva ešte objasniť.
Predpokladá sa, že Enterobacteriaceae zohrávajú úlohu pri kolonizácii B. burgdorferi, pretože jej početnosť negatívne korelovala s abundanciou patogénu [ 55 ]. Tu sme zistili, že protilátkami sprostredkované zacielenie Enterobacteriaceae moduluje mikrobiotu kliešťa a znižuje záťaž B. afzelii v kliešťovi. Tieto výsledky naznačujú, že komenzálne baktérie Enterobacteriaceae môžu mať kľúčovú úlohu pri získavaní patogénov. Keď sme však experimentálne zaviedli E. coli BL21 s B. afzelii do stredného čreva nýmf I. ricinus kapilárnym kŕmením alebo mikroinjekciou, prekvapivo sme zistili, že hladina B. afzelii na týchto nymfách bola nižšia v porovnaní s nymfami, ktoré dostali iba B. afzelii. Tieto výsledky naznačujú, že pozorované zníženie zaťaženia patogénmi nie je spôsobené posunom v množstve Enterobacteriaceae. Je to skôr spôsobené moduláciou mikroflóry (vyčerpaním alebo objavením sa niektorých baktérií) smerom k inému stavu, ako je stav vyvolaný B. afzelii , konkrétne stav odolný voči boréliám nezlučiteľný s vývojom patogénu. Ukázalo sa, že zmena zloženia črevnej mikroflóry stavovcov buď antibiotickou liečbou [ 9 ] alebo pridaním probiotík [ 39 ] môže inhibovať kolonizáciu patogénnych baktérií v čreve. Keď sme definovali tento stav odolný voči infekcii porovnaním taxonomického profilu, hojnosti a objavujúcich sa vlastností mikrobioty kliešťov, zistili sme, že hlavné rozdiely medzi neinfikovaným, patogénnym permisívnym a infekciou odolným stavom spočívali hlavne v kolísaní množstva jedinečných taxónov a vznikajúcich vlastnostiach mikrobiálnych sietí. Emergentné vlastnosti sú jednou zo silných stránok biologických sietí a môžu pomôcť vysvetliť správanie zložitých systémov [ 56 , 57 ]. Zistili sme, že zmeny vyvolané B. afzelii , pokiaľ ide o interagujúce uzly, mikrobiálne-mikrobiálne interakcie a robustnosť mikrobiálnej siete, neboli také odlišné ako zmeny vyvolané E. coli + B. afzelii v porovnaní s neinfikovanou sieťou, keď sú kliešte vystavené imunizovaným myšiam. V sieti E. coli + B. afzelii sa v porovnaní s neinfikovanou alebo B. afzelii zistil najmä počet veľkých okrajov, vyšší počet nových prepojených taxónov a vysoká robustnosť.siete. Keď bola umelo pridaná komenzálna baktéria, zistili sme naopak nižší počet hrán a prepojených taxónov, ako aj nižšiu robustnosť. Tieto výsledky naznačujú, že narušenie mikrobioty rôznymi prostriedkami môže viesť k rôznym stavom odolným voči infekcii. Podobné výsledky boli zistené v mikrobiote kliešťov po narušení vakcínou proti kliešťom, infekciou A. phagocytophilum a antimikrobiálnym peptidom, kde bol zistený vyšší počet asociácií, ale nižšia robustnosť [ 58 ]. Tieto výsledky naznačujú, že kliešťová mikroflóra je vysoko citlivá na poruchy, ktoré viedli k zmenám vo vznikajúcich vlastnostiach mikrobiálnych sietí. Konkrétne sme tu zistili, že narušenie mikroflóry vakcínou proti mikroflóre vyvolalo drastické zmeny v zostave bakteriálnej komunity, ktoré viedli k nevhodnému štádiu na kolonizáciu B. afzelii vo vektore. Je pozoruhodné, že nedávno Narasimhan a kol. [ 59 ] preukázali, že narušenie zloženia mikrobiómu rôznymi stratégiami (nárast I. scapularis v bezmikrobných izolátoroch na generovanie lariev, ktoré neobsahujú žiadne baktérie z prostredia, kŕmenie kliešťami infikovanými B. burgdorferi rezistentnými na gentamicín na myšiach s injekciou gentamicínu, sprostredkované RNAi na I. knockdown statu B. kliešte. Je zaujímavé, že žiadna z metód na narušenie kliešťovej mikroflóry, ktoré sme použili v tejto štúdii (antimikrobiotová vakcína a pridanie komenzálnych baktérií E. coli ), nebola zahrnutá do štúdie Narasimhana et al. [ 59 ]. To naznačuje, že technika použitá na narušenie mikrobioty môže ovplyvniť výsledok kolonizácie patogénom a zdôrazňuje potenciál použitia antimikrobiotickej vakcíny na kontrolu kolonizácie borélií v kliešťoch.
Nakoniec sme zistili, že kliešte kŕmené myšami imunizovanými E. coli BL21 a infikovanými B. afzelii mali významne nižšiu mieru úmrtnosti v porovnaní so skupinou PBS + B. afzelii . Tento výsledok je v kontraste s jednou štúdiou, v ktorej nenašli dôkazy, že infekcia B. afzelii a redukcia mikroflóry lariev (sterilizáciou povrchu vajíčka bielidlom a etanolom) ovplyvnila prežitie kliešťov [ 60 ]. Rozdiely možno vysvetliť rôznymi vývojovými štádiami, v ktorých sa opatrenie vykonalo. V našej štúdii bola úmrtnosť kliešťov meraná v prekrvených larvách, zatiaľ čo v štúdii Hurryho et al. [ 60 ] sa miera prežitia merala u nymf. Predpokladali sme, že zníženie úmrtnosti lariev E. coli + B. afzelii by mohlo byť spôsobené nižšou záťažou B. afzelii v rámci kliešťov, čo by mohlo priaznivo pôsobiť na zdatnosť vektora.
Závery
Zistili sme, že infekcia B. afzelii moduluje mikrobiotu I. ricinus z hľadiska beta diverzity, zloženia, abundancie a mikrobiálneho spoločného výskytu. Široké účinky vyvolané patogénom na kliešťovú mikroflóru sú pravdepodobne výsledkom toho, že patogén vytvára prostredie vhodné na jeho kolonizáciu vo vektore. Narušenie tohto stavu mikrobiómu, ktorý umožňuje infekciu, môže byť alternatívou blokovania kolonizácie patogénom a jeho následného prenosu na nového hostiteľa. Efektívne reťazce infekcie vektormi prenášaných patogénov zahŕňajú kompetentný vektor, infekčný patogén a infekciu permisívny mikrobióm [ 61 ] a nesúlad jednej z týchto zložiek môže mať za následok neschopnosť patogénu účinne kolonizovať črevo vektora a/alebo neschopnosť vektora prenášať patogény [ 61 ]. Tu sme ukázali, že antimikrobiálna vakcína zameraná na Escherichia-Shigella môže formovať mikrobióm I. ricinus smerom k stavu odolnému voči infekcii, posunutím množstva niekoľkých bakteriálnych členov mikrobioty a zvýšením interakcií a robustnosti mikróbov a mikróbov, čo ovplyvnilo celú mikrobiotu kliešťov a viedlo k nižšej záťaži B. afzelii vo vektore. Preto je antimikrobiota vakcína vhodným nástrojom na manipuláciu mikrobiómu do požadovaného stavu a môže byť použitá na kontrolu chorôb prenášaných vektormi.
Doplňujúce informácie
40168_2023_1599_MOESM1_ESM.tif (13,9 MB, tif)Doplnkový súbor 1: Doplnkový obrázok S1. Zmeny v taxonomickom profile mikrobioty kliešťov po infekcii B. afzelii . Graf sopky ukazujúci rozdielnu mikrobiálnu abundanciu v mikrobiote kliešťov z neinfikovaných skupín a skupín B. afzelii . Tyrkysové a fialové bodky predstavujú bakteriálne taxóny, ktorých abundancia v mikrobiote kliešťov zo skupiny B. afzelii v porovnaní s kontrolnou skupinou výrazne klesla, respektíve vzrástla. Teplotné mapy predstavujú početnosť (vyjadrenú ako CLR) všetkých taxónov s významnými rozdielmi medzi neinfikovanou skupinou a skupinou B. afzelii . Taxóny, ktorých abundancia výrazne klesla v skupine B. afzelii, sú označené tyrkysovou farbou. Taxonomická tabuľka použitá na diferenciálnu analýzu abundancie bola získaná zo sekvencií génu 16S rRNA z kliešťov kŕmených neinfikovanými myšami ( n = 10 jednotlivých lariev) a myšiami infikovanými B. afzelii ( n = 10 jednotlivých lariev).
40168_2023_1599_MOESM2_ESM.tif (3,6 MB, tif)Doplnkový súbor 2: Doplnkový obrázok S2. Detekcia proteínov Borrelia . Proteíny borélií sa detegovali westernovým prenosom s použitím séra myší experimentálne infikovaných B. afzelii a imunizovaných živou vakcínou obsahujúcou E. coli BL21 alebo falošnou vakcínou (PBS).
40168_2023_1599_MOESM3_ESM.tif (12 MB, tif)Doplnkový súbor 3: Doplnkový obrázok S3. Zmeny v taxonomickom profile mikroflóry kliešťov po infekcii B. afzelii a imunizácii vakcínou proti mikroflóre. Graf sopky zobrazujúci rozdielnu mikrobiálnu abundanciu v mikrobiote kliešťov zo skupín PBS+ B. afzelii a E. coli + B. afzelii . Zelené a ružové bodky predstavujú bakteriálne taxóny, ktorých početnosť v mikrobiote kliešťov zo skupiny E. coli + B. afzelii v porovnaní so skupinou PBS+ B. afzelii významne klesla, respektíve vzrástla. Teplotné mapy predstavujú početnosť (vyjadrenú ako CLR) všetkých taxónov s významnými rozdielmi medzi skupinami PBS+ B. afzelii a E. coli + B. afzelii . Taxóny, ktorých početnosť v skupine E. coli + B. afzelii výrazne klesla , sú označené zelenou farbou. Taxonomická tabuľka použitá na diferenciálnu analýzu abundancie bola získaná zo sekvencií génu 16S rRNA z kliešťov kŕmených PBS+ myšiam B. afzelii ( n = 10 jednotlivých lariev) a myšiam E. coli + B. afzelii ( n = 8 jednotlivých lariev).
40168_2023_1599_MOESM4_ESM.tif (4,8 MB, tif)Doplnkový súbor 4: Doplnkový obrázok S4. Vplyv pridania komenzálnej baktérie na alfa a beta diverzitu kliešťovej mikroflóry. (a) Fylogenetická diverzita Faith a (b) Pielouove indexy rovnomernosti sa použili na meranie bohatosti a rovnomernosti mikrobioty kliešťov, ktoré dostali B. afzelii alebo E. coli+B. afzelii kapilárnym kŕmením alebo injekciou do análnych pórov (Kruskal-Wallis, p < 0,05 ). Beta diverzita kliešťovej mikroflóry sa analyzovala pomocou (c) Jaccard a (d) vážených Unifrac indexov na meranie podobnosti medzi bakteriálnymi komunitami v mikrobiote kliešťov, ktoré dostali B. afzelii alebo E. coli+B. afzelii kapilárnym kŕmením alebo injekciou análnych pórov za rôznych experimentálnych podmienok (PERMANOVA, p < 0,05 ).
40168_2023_1599_MOESM5_ESM.tif (12,7 MB, tif)Doplnkový súbor 5: Doplnkový obrázok S5. Detekcia Enterobacteriaceae pomocou PCR. Reprezentatívne obrázky gélu elektroforézy ukazujúce pásy zodpovedajúce génu 16S rRNA pre Enterobacteriaceae. Rôzne panely predstavujú rôzne experimenty: Kliešte dostali B. afzelii alebo E. coli + B. afzelii kapilárnym kŕmením a inkubovali sa 6 hodín po kŕmení (horný panel) alebo mikroinjekciou a inkubovali sa 6 hodín po injekcii (spodný panel). Každý pruh predstavuje inú značku zo skupín. Na pozitívnu kontrolu bola použitá DNA extrahovaná z kultúry E. coli BL21.
40168_2023_1599_MOESM6_ESM.tif (4,1 MB, tif)Doplnkový súbor 6: Doplnkový obrázok S6. Zhluková analýza rôznych vzoriek kliešťovej mikrobioty. Dendrogram založený na Wardovej metóde zhlukovania vzoriek kliešťovej mikroflóry z neinfikovaných skupín B. afzelii , PBS+ B. afzelii a E. coli + B. afzelii .
40168_2023_1599_MOESM7_ESM.tif (11,8 MB, tif)Doplnkový súbor 7: Doplnkový obrázok S7. Vplyv pridania komenzálnej baktérie na vznikajúce vlastnosti I. ricinus mikrobiota. Vennov diagram znázorňujúci (a) bežné a jedinečné bakteriálne taxóny medzi kliešťovou mikroflórou, ktorá dostala B. afzelii alebo E. coli+B. afzelii kapilárnym kŕmením a injekciou análnych pórov (b) zdieľané a jedinečné taxóny, ktorých početnosť sa významne zmenila medzi porovnaním B. afzelii vs. E. coli + B. afzelii v skupinách s kapilárnym kŕmením a injekciou do análnych pórov, (c) Dendrogram zhlukovania vzoriek kliešťovej mikroflóry v rôznych experimentálnych podmienkach, (d) nájdený spoločný diagram mikrobiálnych uzlíkov zo spoločných a Vendencových nodov podmienky. Bodový graf zobrazujúci priemer pozorovaných znakov oproti počtu (e) uzlov a (f) okrajov nájdených v sieťach mikrobiálneho spoločného výskytu a (g) bodový graf zobrazujúci stratu konektivity, keď sa zo siete mikrobiálneho spoločného výskytu odstráni 5 až 7 % uzlov.
40168_2023_1599_MOESM8_ESM.csv (1,6 MB, csv)Doplnkový súbor 8: Doplnková tabuľka S1. Bakteriálne taxóny nájdené ako kontaminanty v súboroch údajov o sekvenovaní génov 16S rRNA z kliešťov kŕmených myšami v rôznych experimentálnych podmienkach. Kontaminanty boli štatisticky identifikované (TRUE) a odstránené zo súborov údajov o sekvenovaní génov 16S rRNA pomocou balíka decontam R
40168_2023_1599_MOESM9_ESM.xlsx (17,7 kB, xlsx)Doplnkový súbor 9: Doplnková tabuľka S2. Jaccardove indexy miestnej centrality merajú na porovnanie medzi neinfikovanými skupinami a skupinami B. afzelii . Jaccardove indexy pre každú z mier lokálnej centrálnosti (tj stupeň, centrálnosť medziľahlosti, centrálnosť blízkosti, centrálnosť vlastného vektora a uzlové taxóny) súborov väčšiny centrálnych uzlov na porovnávanie párových sietí. Boli sčítané dve p-hodnoty, P(J ≤ j) a P(J ≥ j), pre každý Jaccardov index.
40168_2023_1599_MOESM10_ESM.xlsx (17,5 kB, xlsx)Doplnkový súbor 10: Doplnková tabuľka S3. Spoločné a jedinečné susedné uzly taxónu Escherichia-Shigella medzi neinfikovanou skupinou a skupinou B. afzelii .
40168_2023_1599_MOESM11_ESM.xlsx (18,3 KB, xlsx)Doplnkový súbor 11: Doplnková tabuľka S4. Tkanivá myší pozitívne na B. afzelii v PBS+ B. afzelii a E . coli+B. afzelii. Srdce, koža a pravý členkový kĺb boli testované na DNA B. afzelii a pozitívne tkanivá boli uvedené v paneli a a b pre PBS+ B. afzelii a E . coli+B. afzelii, resp . Srdce a koža boli testované na B. afzelii RNA a pozitívne tkanivá boli uvedené v paneli c a d pre PBS+ B. afzelii a E . coli+B. afzelii, resp.
40168_2023_1599_MOESM12_ESM.xlsx (18,5 kB, xlsx)Doplnkový súbor 12: Doplnková tabuľka S5. Jaccardove indexy meraní lokálnej centrality na porovnanie medzi PBS+ B. afzelii vs. E. coli+B. afzelii skupiny. Jaccardove indexy pre každú z mier lokálnej centrálnosti (tj stupeň, centrálnosť medziľahlosti, centrálnosť blízkosti, centrálnosť vlastného vektora a uzlové taxóny) súborov väčšiny centrálnych uzlov na porovnávanie párových sietí. Dve p-hodnoty, P(J ≤ j) a P(J ≥ j), pre každý Jaccardov index sa pridali
40168_2023_1599_MOESM13_ESM.xlsx (16,7 kB, xlsx)Doplnkový súbor 13: Doplnková tabuľka S6. Spoločné a jedinečné susedné uzly taxónu Escherichia-Shigella medzi PBS+ B. afzelii a E. coli+B. afzelii skupiny.
40168_2023_1599_MOESM14_ESM.csv (1,8 MB, csv)Doplnkový súbor 14: Doplnková tabuľka S7. Bakteriálne taxóny nachádzajúce sa ako kontaminanty v súboroch údajov o sekvenovaní génu 16S rRNA z kliešťov vystavených B. afzelii alebo E. coli+B. afzelii kapilárnym kŕmením alebo mikroinjekciou do análnych pórov. Kontaminanty boli štatisticky identifikované (TRUE) a odstránené z dátových súborov sekvenovania génu 16S rRNA pomocou balíka decontam R.
Poďakovanie
Ďakujeme členom nášho laboratória za poučné diskusie o experimentoch a výsledkoch prezentovaných v tomto rukopise.
Príspevky autorov
ACC konceptualizoval a koncipoval myšlienku. AWC, ACC, LMH, DO, ROMR a LS koncipovali experimenty. Experimenty vykonali AWC, LMH, AM, SP, SR, LS a AFS. ROMR a RS dohliadali na experimenty s infekciou Borrelia. AWC, ACC, JA, AM a DO analyzovali a/alebo interpretovali výsledky. ACC, SM, LS a VP poskytli zdroje a vstupy do experimentov. AWC a ACC vypracovali prvú verziu rukopisu. AWC, LMH, AM, ROMR, RS, SP, SR, AFS, SM, VP, JA, LS, DO a ACC rukopis posúdili, upravili a schválili.
Financovanie
ACC, LS a SM (UMR BIPAR) boli podporené francúzskym vládnym programom Investissement d’Avenir, Laboratoire d’Excellence „Integrovaná biológia vznikajúcich infekčných chorôb“ (č. grantu ANR-10-LABX-62-IBEID). AWC podporil Programa Nacional de Becas de Postgrado en el Exterior “Don Carlos Antonio López” (grant č. 205/2018). AM je podporovaný grantom „Collectivité de Corse“: „Formations superieures“ (SGCE-RAPPORT č. 0300). RS podporilo Ministerstvo zdravotníctva ČR, grant č. NU20-05-00396 a grantom Grantovej agentúry Českej republiky č. 22-30920S.
Dostupnosť údajov a materiálov
Súbory údajov prezentované v tejto štúdii možno nájsť v online úložiskách. Názvy repozitárov/úložísk a prístupové čísla (čísla) nájdete nižšie: https://www.ncbi.nlm.nih.gov/sra , PRJNA870490.
vyhlásenia
Etický súhlas a súhlas s účasťou
Experimenty in vivo sa uskutočnili v zariadení pre zvieratá Laboratória pre zdravie zvierat Francúzskej agentúry pre bezpečnosť potravín, životného prostredia a pri práci (ANSES), Maisons-Alfort, Francúzsko, podľa francúzskych a medzinárodných hlavných zásad pre biomedicínsky výskum zahŕňajúci zvieratá (2012). Postupy preverila a schválila Etická komisia (ComEth, Anses/ENVA/UPEC) s číslom povolenia na pokusy na zvieratách E 94 046 08.
Súhlas so zverejnením
Neuplatňuje sa.
Konkurenčné záujmy
Autori nedeklarujú žiadne konkurenčné záujmy.
Poznámky pod čiarou
Poznámka vydavateľa
Springer Nature zostáva neutrálny, pokiaľ ide o jurisdikčné nároky v publikovaných mapách a inštitucionálnych afiliáciách.
Referencie
- 1.Madison-Antenucci S, Kramer LD, Gebhardt LL, Kauffman E. Vznikajúce choroby prenášané kliešťami. Clin Microbiol Rev. 2020;33:e00083–e118. doi: 10.1128/CMR.00083-18. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 2.Radolf JD, Caimano MJ, Stevenson B, Hu LT. O kliešťoch, myšiach a mužoch: pochopenie dvojhostiteľského životného štýlu spirochétov Lymskej choroby. Nat Rev Microbiol. 2012;10:87–99. doi: 10.1038/nrmicro2714. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 3.Kurokawa C, Lynn GE, Pedra JHF, Pal U, Narasimhan S, Fikrig E. Interakcie medzi Borrelia burgdorferi a kliešťami. Nat Rev Microbiol. 2020;18:587–600. doi: 10.1038/s41579-020-0400-5. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 4.Barbour AG, Fish D. Biologický a sociálny fenomén lymskej boreliózy. Veda. 1993;260:1610–1616. doi: 10.1126/science.8503006. [ DOI ] [ PubMed ] [ Študovňa Google ]
- 5.Richter D, Klug B, Spielman A, Matuschka FR. Adaptácia rôznych spirochét Lymskej choroby v prirodzenom hostiteľovi rezervoáru hlodavcov. Infect Immun. 2004;72:2442. doi: 10.1128/IAI.72.4.2442-2444.2004. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 6.Cirimotich CM, Ramirez JL, Dimopoulos G. Kompetencia vektora hmyzu v tvare prirodzenej mikrobioty pre ľudské patogény. Bunkový hostiteľský mikrób. 2011;10:307–310. doi: 10.1016/j.chom.2011.09.006. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 7.Dong Y, Manfredini F, Dimopoulos G. Dôsledok mikrobioty stredného čreva komára v obrane proti parazitom malárie. PLoS Pathog. 2009;5:e1000423. doi: 10.1371/journal.ppat.1000423. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 8.Xi Z, Ramirez JL, Dimopoulos G. Cesta Aedes aegypti Toll kontroluje infekciu vírusom dengue. PLoS Pathog. 2008;4:e1000098. doi: 10.1371/journal.ppat.1000098. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 9.Narasimhan S, Rajeevan N, Liu L, Zhao YO, Heisig J, Pan J a kol. Črevná mikroflóra kliešťového vektora Ixodes scapularis moduluje kolonizáciu spirochéty Lymskej choroby. Bunkový hostiteľský mikrób. 2014;15:58–71. doi: 10.1016/j.chom.2013.12.001. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 10.Bando H, Okado K, Guelbeogo WM, Badolo A, Aonuma H, Nelson B a kol. Vnútrošpecifická diverzita Serratia marcescens v strednom čreve komára Anopheles definuje prenosovú kapacitu Plasmodium. Sci Rep. 2013;3:1641. doi: 10.1038/srep01641. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 11.Gall CA, Reif KE, Scoles GA, Mason KL, Mousel M, Noh SM a kol. Bakteriálny mikrobióm kliešťov Dermacentor andersoni ovplyvňuje citlivosť patogénov. ISME J. 2016;10:1846–1855. doi: 10.1038/ismej.2015.266. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 12.Landesman WJ, Mulder K, Fredericks LP, Allan BF. Krížová analýza mikrobiálnych spoločenstiev Ixodes scapularis v nymfálnom štádiu vo vzťahu k infekcii a záťaži Borrelia burgdorferi. FEMS Microbiol Ecol. 2019;95(12):fiz167. 10.1093/femsec/fiz167. [ DOI ] [ bezplatný článok PMC ] [ PubMed ]
- 13.Mateos-Hernández L, Obregón D, Maye J, Bornères J, Versille N, de la Fuente J, et al. Vakcína proti kliešťovej mikroflóre ovplyvňuje výkonnosť Ixodes ricinus počas kŕmenia. Vakcíny (Bazilej) 2020; 8:702. doi: 10 3390/vakcíny8040702. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 14.Mateos-Hernández L, Obregón D, Wu-Chuang A, Maye J, Bornères J, Versillé N a kol. Antimikrobiálne vakcíny modulujú mikrobióm kliešťov spôsobom špecifickým pre taxóny. Front Immunol. 2021;12:704621. doi: 10.3389/fimmu.2021.704621. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 15.Aželytė J, Wu-Chuang A, Žiegytė R, Platonova E, Mateos-Hernandez L, Maye J a kol. Antimikrobiotová vakcína znižuje infekciu vtáčou maláriou v rámci vektorov komárov. Front Immunol. 2022;13:841835. doi: 10.3389/fimmu.2022.841835. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 16.Schwan TG, Simpson WJ, Rosa PA. Laboratórne potvrdenie lymskej boreliózy. Can J Infect Dis. 1991;2:64–69. doi: 10.1155/1991/637201. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 17.Mateos-Hernández L, Rakotobe S, Defaye B, Cabezas-Cruz A, Šimo L. Model založený na kapsulách na napadnutie štádiami nezrelých tvrdých kliešťov na laboratórnych myšiach. J Vis Exp. 2020 doi: 10.3791/61430. [ DOI ] [ PubMed ] [ Študovňa Google ]
- 18.Castillo M, Martín-Orúe SM, Manzanilla EG, Badiola I, Martín M, Gasa J. Kvantifikácia celkových populácií baktérií, enterobaktérií a laktobacilov v tráve ošípaných pomocou PCR v reálnom čase. Vet Microbiol. 2006;114:165–170. doi: 10.1016/j.vetmic.2005.11.055. [ DOI ] [ PubMed ] [ Študovňa Google ]
- 19.Davis NM, Proctor DM, Holmes SP, Relman DA, Callahan BJ. Jednoduchá štatistická identifikácia a odstránenie kontaminujúcich sekvencií z markerových génov a metagenomických údajov. Mikrobióm. 2018;6:226. doi: 10.1186/s40168-018-0605-2. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 20.Bolyen E, Rideout JR, Dillon MR, Bokulich NA, Abnet M, Asnicar F a kol. Reprodukovateľná, interaktívna, škálovateľná a rozšíriteľná veda o mikrobiómoch pomocou QIIME 2. Nat Biotechnol. 2019;37:852–857. doi: 10.1038/s41587-019-0209-9. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 21.Callahan BJ, McMurdie PJ, Rosen MJ, Han AW, Johnson AJA, Holmes SP. DADA2: Vzorový záver s vysokým rozlíšením z údajov amplikónu Illumina. Metódy Nat. 2016;13:581–583. doi: 10.1038/nmeth.3869. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 22.Bokulich NA, Kaehler BD, Rideout JR, Dillon M, Bolyen E, Knight R a kol. Optimalizácia taxonomickej klasifikácie amplikónových sekvencií markerového génu pomocou doplnku QIIME 2 q2-feature-classifier. Mikrobióm. 2018;6:90. doi: 10.1186/s40168-018-0470-z. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 23.Yarza P, Yilmaz P, Pruesse E, Glöckner FO, Ludwig W, Schleifer KH a kol. Zjednotenie klasifikácie kultivovaných a nekultivovaných baktérií a archaea pomocou génových sekvencií 16S rRNA. Nat Rev Microbiol. 2014;12:635–645. doi: 10.1038/nrmicro3330. [ DOI ] [ PubMed ] [ Študovňa Google ]
- 24.Margos G, Gofton A, Wibberg D, Dangel A, Marosevic D, Loh SM a kol. Rod Borrelia reloaded. PLoS ONE. 2018;13:e0208432. doi: 10.1371/journal.pone.0208432. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 25.Adeolu M, Gupta RS. Fylogenomický a molekulárny markerový návrh na rozdelenie rodu Borrelia do dvoch rodov: emendovaný rod Borrelia obsahujúci iba členov recidivujúcej horúčky Borrelia a rod Borreliella gen. nov. obsahujúcich členov boréliovej lymskej choroby (komplex Borrelia burgdorferi sensu lato) Antonie van Leeuwenhoek, Int J Gen Mol Microbiol. 2014;105:1049–72. doi: 10.1007/s10482-014-0164-x. [ DOI ] [ PubMed ] [ Študovňa Google ]
- 26. Gupta RS. Rozlíšenie medzi Borrelia a Borreliella je silnejšie podporované molekulárnymi a fenotypovými charakteristikami ako všetky ostatné susedné prokaryotické rody: Reakcia na Margos‘ et al. „Reloaded rod Borrelia“ (PLoS ONE 13(12): e0208432). PLoS One. 2019;14(8):e0221397. 10.1371/journal.pone.0221397. [ DOI ] [ bezplatný článok PMC ] [ PubMed ]
- 27.Friedman J, Alm EJ. Odvodzovanie korelačných sietí z údajov genomického prieskumu. PLoS Comput Biol. 2012;8:e1002687. doi: 10.1371/journal.pcbi.1002687. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 28.Tím RStudio. RStudio: Integrovaný vývoj pre R. Boston: RStudio, PBC; 2020. http://www.rstudio.com/ .
- 29. Bastian M, Heymann S, Jacomy, M. Gephi: Softvér s otvoreným zdrojom na skúmanie a manipuláciu sietí. Zborník z medzinárodnej konferencie AAAI o webe a sociálnych médiách. 2009;3(1):361–62. 10.1609/icwsm.v3i1.13937.
- 30.Lhomme S. NetSwan: Analýza silných a slabých stránok siete. Verzia R Pack. 2015. https://rdrr.io/cran/NetSwan/ . Prístup v septembri 2022.
- 31.Peschel S, Müller CL, von Mutius E, Boulesteix AL, Depner M. NetCoMi: konštrukcia siete a porovnanie údajov o mikrobiómoch v R. Brief Bioinform. 2021;22(4):bbaa290. 10.1093/bib/bbaa290. [ DOI ] [ bezplatný článok PMC ] [ PubMed ]
- 32.Fernandes AD, Reid JN, Macklaim JM, McMurrough TA, Edgell DR, Gloor GB. Zjednotenie analýzy vysokovýkonných sekvenčných súborov údajov: charakterizácia RNA-seq, sekvenovanie génov 16S rRNA a selektívne rastové experimenty analýzou kompozičných údajov. Mikrobióm. 2014;2:15. doi: 10.1186/2049-2618-2-15. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 33.Love MI, Huber W, Anders S. Moderovaný odhad násobnej zmeny a disperzie pre údaje RNA-seq s DESeq2. Genome Biol. 2014;15:550. doi: 10.1186/s13059-014-0550-8. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 34.Oksanen J, Simpson GL, Blanchet FG, Kindt R, Legendre P, Minchin PR a kol. Vegan: Balíček ekológie komunity. R Packag verzia 26-0. 2021. https://CRAN.R-project.org/package=vegan . Prístupné v auguste 2023.
- 35.Fawcett PT, Rose CD, Gibney KM. Porovnávacie vyhodnotenie adsorpcie s E. coli na testoch ELISA na lymskú boreliózu. J Rheumatol. 1995;22(4):684–688. [ PubMed ] [ Študovňa Google ]
- 36.Bruckbauer HR, Preac-Mursic V, Fuchs R, Wilske B. Krížovo reaktívne proteíny Borrelia burgdorferi. Eur J Clin Microbiol Infect Dis. 1992;11(3):224–232. doi: 10.1007/BF02098084. [ DOI ] [ PubMed ] [ Študovňa Google ]
- 37.Liu Y, Wang J, Wu C. Modulácia črevnej mikroflóry a imunitného systému probiotikami, prebiotikami a postbiotikami. Predné Nutr. 2022;8:634897. doi: 10.3389/fnut.2021.634897. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 38.Everard A, Matamoros S, Geurts L, Delzenne NM, Cani PD. Podávanie Saccharomyces boulardii mení črevnú mikrobiotu a znižuje steatózu pečene, zápal nízkeho stupňa a tukovú hmotu u obéznych a diabetických db/db myší typu 2. MBio. 2014;5:e01011–e1014. doi: 10.1128/mBio.01011-14. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 39.Wang X, Zhang P, Zhang X. Probiotiká regulujú črevnú mikroflóru: účinná metóda na zlepšenie imunity. Molekuly. 2021;26:6076. doi: 10,3390/molekuly26196076. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 40.Tuo Y, Song X, Song Y, Liu W, Tang Y, Gao Y a kol. Skríning probiotík z kmeňov Lactobacillus podľa ich schopností inhibovať adhéziu patogénov a indukciu prozápalového cytokínu IL-8. J Dairy Sci. 2018;101:4822–4829. doi: 10.3168/jds.2017-13654. [ DOI ] [ PubMed ] [ Študovňa Google ]
- 41.Surendran Nair M, Amalaradjou MA, Venkitanarayanan K. Antivirulentné vlastnosti probiotík v boji proti mikrobiálnej patogenéze. Adv Appl Microbiol. 2017 doi: 10.1016/bs.aambs.2016.12.001. [ DOI ] [ PubMed ] [ Študovňa Google ]
- 42.Fang K, Jin X, Hong SH. Probiotikum Escherichia coli inhibuje tvorbu biofilmu patogénnej E. coli prostredníctvom extracelulárnej aktivity DegP. Sci Rep. 2018;8(1):4939. 10.1038/s41598-018-23180-1. [ DOI ] [ bezplatný článok PMC ] [ PubMed ]
- 43.Študoval FW, Moffatt BA. Použitie bakteriofágovej T7 RNA polymerázy na riadenie selektívnej vysokoúrovňovej expresie klonovaných génov. J Mol Biol. 1986;189:113–130. doi: 10.1016/0022-2836(86)90385-2. [ DOI ] [ PubMed ] [ Študovňa Google ]
- 44.Jeong H, Kim HJ, Lee SJ. Kompletná genómová sekvencia Escherichia coli kmeňa BL21. Oznámenie genómu. 2015;3:134–149. doi: 10.1128/genómA.00134-15. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 45.Narasimhan S, Swei A, Abouneameh S, Pal U, Pedra JHF, Fikrig E. Zápasenie s mikrobiómom kliešťov. Trendy Parasitol. 2021;37:722. doi: 10.1016/j.pt.2021.04.004. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 46.Wu-Chuang A, Hodžić A, Mateos-Hernández L, Estrada-Peña A, Obregon D, Cabezas-Cruz A. Aktuálne diskusie a pokroky vo výskume mikrobiómov kliešťov. Curr Res Parasitol Vector-Borne Dis. 2021;1:100036. doi: 10.1016/j.crpvbd.2021.100036. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 47.Dennison NJ, Jupatanakul N, Dimopoulos G. Mikrobiota komárov ovplyvňuje kompetenciu vektorov pre ľudské patogény. Curr Opin Insect Sci. 2014;3:6–13. doi: 10.1016/j.cois.2014.07.004. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 48.Hamilton PT, Maluenda E, Sarr A, Belli A, Hurry G, Duron O a kol. Infekcia Borrelia afzelii v hostiteľovi hlodavcov má dramatické účinky na bakteriálny mikrobióm kliešťov Ixodes ricinus. Appl Environ Microbiol. 2021;87:e0064121. doi: 10.1128/AEM.00641-21. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 49.Chauhan G, McClure J, Hekman J, Marsh PW, Bailey JA, Daniels RF a kol. Kombinácia občianskej vedy a genomiky na skúmanie kliešťov, patogénov a komenzálneho mikrobiómu pri rozlíšení jedného kliešťa. Predná Genet. 2020;10:1322. doi: 10.3389/fgene.2019.01322. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 50.Lejal E, Chiquet J, Aubert J, Robin S, Estrada-Peña A, Rue O a kol. Časové vzorce v mikrobiálnych spoločenstvách Ixodes ricinus: pohľad na interakcie mikróbov prenášaných kliešťami. Mikrobióm. 2021;9:153. doi: 10.1186/s40168-021-01051-8. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 51.Narasimhan S, Schuijt TJ, Abraham NM, Rajeevan N, Coumou J, Graham M a kol. Modulácia prostredia kliešťa vylučovaným kliešťovým proteínom podporuje kolonizáciu Borrelia burgdorferi. Nat Commun. 2017;8:184. doi: 10.1038/s41467-017-00208-0. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 52.Abraham NM, Liu L, Jutras BL, Yadav AK, Narasimhan S, Gopalakrishnan V a kol. Patogénom sprostredkovaná manipulácia s mikrobiotou článkonožcov na podporu infekcie. Proc Natl Acad Sci US A. 2017;114:E781–E790. doi: 10.1073/pnas.1613422114. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 53.Wu-Chuang A, Obregon D, Mateos-Hernández L, Cabezas-Cruz A. Vakcíny proti kliešťom: ako to vlastne môže fungovať? Biológia. 2022;77:1555–1562. doi: 10.1007/s11756-021-00818-6. [ DOI ] [ Študovňa Google ]
- 54.Rana VS, Kitsou C, Dutta S, Ronzetti MH, Zhang M, Bernard Q a kol. Signalizácia Dome1-JAK-STAT medzi parazitom a hostiteľom integruje vektorovú imunitu a vývoj. Veda. 2023;379(6628):eabl3837. doi: 10.1126/science.abl3837. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 55.Ross BD, Hayes B, Radey MC, Lee X, Josek T, Bjork J a kol. Ixodes scapularis nemá stabilný mikrobióm stredného čreva. ISME J. 2018;12:2596–2607. doi: 10.1038/s41396-018-0161-6. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 56.Aderem A. Systémová biológia: Jej prax a výzvy. Cell. 2005;121:511–513. doi: 10.1016/j.cell.2005.04.020. [ DOI ] [ PubMed ] [ Študovňa Google ]
- 57.Röttjers L, Faust K. Od chĺpkov k hypotézam – biologické poznatky z mikrobiálnych sietí. FEMS Microbiol Rev. 2018;42:761–780. doi: 10.1093/femsre/fuy030. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 58.Estrada-Peña A, Cabezas-Cruz A, Obregón D. Odolnosť črevného mikrobiómu kliešťa voči vakcínam proti kliešťom, infekcii patogénom a antimikrobiálnym peptidom. Patogény. 2020; 9:309. doi: 10.3390/patogény9040309. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 59.Narasimhan S, Rajeevan N, Graham M, Wu MJ, DePonte K, Marion S a kol. Prenos kliešťov Borrelia burgdorferi na myšieho hostiteľa nie je ovplyvnený mikrobiotou stredného čreva získanou z prostredia. Mikrobióm. 2022;10:173. doi: 10.1186/s40168-022-01378-w. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 60.Hurry G, Maluenda E, Sarr A, Belli A, Hamilton PT, Duron O a kol. Infekcia Borrelia afzelii a manipulácia s mikrobiotou na povrchu vajíčka nemajú žiadny vplyv na kondíciu nedospelých kliešťov Ixodes ricinus. Sci Reports. 2021;11:10686. doi: 10.1038/s41598-021-90177-8. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
- 61.Maitre A, Wu-Chuang A, Aželytė J, Palinauskas V, Mateos-Hernandez L, Obregon D a kol. Manipulácia s vektorovou mikrobiotou hostiteľskými protilátkami: zabudnutá stratégia vývoja vakcín blokujúcich prenos. vektory parazitov. 2022;15:4. doi: 10.1186/s13071-021-05122-5. [ DOI ] [ bezplatný článok PMC ] [ PubMed ] [ Google Scholar ]
Pridružené údaje
Táto časť zhromažďuje všetky citácie údajov, vyhlásenia o dostupnosti údajov alebo doplňujúce materiály zahrnuté v tomto článku.
Doplnkové materiály
40168_2023_1599_MOESM1_ESM.tif (13,9 MB, tif)Doplnkový súbor 1: Doplnkový obrázok S1. Zmeny v taxonomickom profile mikrobioty kliešťov po infekcii B. afzelii . Graf sopky ukazujúci rozdielnu mikrobiálnu abundanciu v mikrobiote kliešťov z neinfikovaných skupín a skupín B. afzelii . Tyrkysové a fialové bodky predstavujú bakteriálne taxóny, ktorých abundancia v mikrobiote kliešťov zo skupiny B. afzelii v porovnaní s kontrolnou skupinou výrazne klesla, respektíve vzrástla. Teplotné mapy predstavujú početnosť (vyjadrenú ako CLR) všetkých taxónov s významnými rozdielmi medzi neinfikovanou skupinou a skupinou B. afzelii . Taxóny, ktorých abundancia výrazne klesla v skupine B. afzelii, sú označené tyrkysovou farbou. Taxonomická tabuľka použitá na diferenciálnu analýzu abundancie bola získaná zo sekvencií génu 16S rRNA z kliešťov kŕmených neinfikovanými myšami ( n = 10 jednotlivých lariev) a myšiami infikovanými B. afzelii ( n = 10 jednotlivých lariev).
40168_2023_1599_MOESM2_ESM.tif (3,6 MB, tif)Doplnkový súbor 2: Doplnkový obrázok S2. Detekcia proteínov Borrelia . Proteíny borélií sa detegovali westernovým prenosom s použitím séra myší experimentálne infikovaných B. afzelii a imunizovaných živou vakcínou obsahujúcou E. coli BL21 alebo falošnou vakcínou (PBS).
40168_2023_1599_MOESM3_ESM.tif (12 MB, tif)Doplnkový súbor 3: Doplnkový obrázok S3. Zmeny v taxonomickom profile mikroflóry kliešťov po infekcii B. afzelii a imunizácii vakcínou proti mikroflóre. Graf sopky zobrazujúci rozdielnu mikrobiálnu abundanciu v mikrobiote kliešťov zo skupín PBS+ B. afzelii a E. coli + B. afzelii . Zelené a ružové bodky predstavujú bakteriálne taxóny, ktorých početnosť v mikrobiote kliešťov zo skupiny E. coli + B. afzelii v porovnaní so skupinou PBS+ B. afzelii významne klesla, respektíve vzrástla. Teplotné mapy predstavujú početnosť (vyjadrenú ako CLR) všetkých taxónov s významnými rozdielmi medzi skupinami PBS+ B. afzelii a E. coli + B. afzelii . Taxóny, ktorých početnosť v skupine E. coli + B. afzelii výrazne klesla , sú označené zelenou farbou. Taxonomická tabuľka použitá na diferenciálnu analýzu abundancie bola získaná zo sekvencií génu 16S rRNA z kliešťov kŕmených PBS+ myšiam B. afzelii ( n = 10 jednotlivých lariev) a myšiam E. coli + B. afzelii ( n = 8 jednotlivých lariev).
40168_2023_1599_MOESM4_ESM.tif (4,8 MB, tif)Doplnkový súbor 4: Doplnkový obrázok S4. Vplyv pridania komenzálnej baktérie na alfa a beta diverzitu kliešťovej mikroflóry. (a) Fylogenetická diverzita Faith a (b) Pielouove indexy rovnomernosti sa použili na meranie bohatosti a rovnomernosti mikrobioty kliešťov, ktoré dostali B. afzelii alebo E. coli+B. afzelii kapilárnym kŕmením alebo injekciou do análnych pórov (Kruskal-Wallis, p < 0,05 ). Beta diverzita kliešťovej mikroflóry sa analyzovala pomocou (c) Jaccard a (d) vážených Unifrac indexov na meranie podobnosti medzi bakteriálnymi komunitami v mikrobiote kliešťov, ktoré dostali B. afzelii alebo E. coli+B. afzelii kapilárnym kŕmením alebo injekciou análnych pórov za rôznych experimentálnych podmienok (PERMANOVA, p < 0,05 ).
40168_2023_1599_MOESM5_ESM.tif (12,7 MB, tif)Doplnkový súbor 5: Doplnkový obrázok S5. Detekcia Enterobacteriaceae pomocou PCR. Reprezentatívne obrázky gélu elektroforézy ukazujúce pásy zodpovedajúce génu 16S rRNA pre Enterobacteriaceae. Rôzne panely predstavujú rôzne experimenty: Kliešte dostali B. afzelii alebo E. coli + B. afzelii kapilárnym kŕmením a inkubovali sa 6 hodín po kŕmení (horný panel) alebo mikroinjekciou a inkubovali sa 6 hodín po injekcii (spodný panel). Každý pruh predstavuje inú značku zo skupín. Na pozitívnu kontrolu bola použitá DNA extrahovaná z kultúry E. coli BL21.
40168_2023_1599_MOESM6_ESM.tif (4,1 MB, tif)Doplnkový súbor 6: Doplnkový obrázok S6. Zhluková analýza rôznych vzoriek kliešťovej mikrobioty. Dendrogram založený na Wardovej metóde zhlukovania vzoriek kliešťovej mikroflóry z neinfikovaných skupín B. afzelii , PBS+ B. afzelii a E. coli + B. afzelii .
40168_2023_1599_MOESM7_ESM.tif (11,8 MB, tif)Doplnkový súbor 7: Doplnkový obrázok S7. Vplyv pridania komenzálnej baktérie na vznikajúce vlastnosti I. ricinus mikrobiota. Vennov diagram znázorňujúci (a) bežné a jedinečné bakteriálne taxóny medzi kliešťovou mikroflórou, ktorá dostala B. afzelii alebo E. coli+B. afzelii kapilárnym kŕmením a injekciou análnych pórov (b) zdieľané a jedinečné taxóny, ktorých početnosť sa významne zmenila medzi porovnaním B. afzelii vs. E. coli + B. afzelii v skupinách s kapilárnym kŕmením a injekciou do análnych pórov, (c) Dendrogram zhlukovania vzoriek kliešťovej mikroflóry v rôznych experimentálnych podmienkach, (d) nájdený spoločný diagram mikrobiálnych uzlíkov zo spoločných a Vendencových nodov podmienky. Bodový graf zobrazujúci priemer pozorovaných znakov oproti počtu (e) uzlov a (f) okrajov nájdených v sieťach mikrobiálneho spoločného výskytu a (g) bodový graf zobrazujúci stratu konektivity, keď sa zo siete mikrobiálneho spoločného výskytu odstráni 5 až 7 % uzlov.
40168_2023_1599_MOESM8_ESM.csv (1,6 MB, csv)Doplnkový súbor 8: Doplnková tabuľka S1. Bakteriálne taxóny nájdené ako kontaminanty v súboroch údajov o sekvenovaní génov 16S rRNA z kliešťov kŕmených myšami v rôznych experimentálnych podmienkach. Kontaminanty boli štatisticky identifikované (TRUE) a odstránené zo súborov údajov o sekvenovaní génov 16S rRNA pomocou balíka decontam R
40168_2023_1599_MOESM9_ESM.xlsx (17,7 kB, xlsx)Doplnkový súbor 9: Doplnková tabuľka S2. Jaccardove indexy miestnej centrality merajú na porovnanie medzi neinfikovanými skupinami a skupinami B. afzelii . Jaccardove indexy pre každú z mier lokálnej centrálnosti (tj stupeň, centrálnosť medziľahlosti, centrálnosť blízkosti, centrálnosť vlastného vektora a uzlové taxóny) súborov väčšiny centrálnych uzlov na porovnávanie párových sietí. Boli sčítané dve p-hodnoty, P(J ≤ j) a P(J ≥ j), pre každý Jaccardov index.
40168_2023_1599_MOESM10_ESM.xlsx (17,5 kB, xlsx)Doplnkový súbor 10: Doplnková tabuľka S3. Spoločné a jedinečné susedné uzly taxónu Escherichia-Shigella medzi neinfikovanou skupinou a skupinou B. afzelii .
40168_2023_1599_MOESM11_ESM.xlsx (18,3 KB, xlsx)Doplnkový súbor 11: Doplnková tabuľka S4. Tkanivá myší pozitívne na B. afzelii v PBS+ B. afzelii a E . coli+B. afzelii. Srdce, koža a pravý členkový kĺb boli testované na DNA B. afzelii a pozitívne tkanivá boli uvedené v paneli a a b pre PBS+ B. afzelii a E . coli+B. afzelii, resp . Srdce a koža boli testované na B. afzelii RNA a pozitívne tkanivá boli uvedené v paneli c a d pre PBS+ B. afzelii a E . coli+B. afzelii, resp.
40168_2023_1599_MOESM12_ESM.xlsx (18,5 kB, xlsx)Doplnkový súbor 12: Doplnková tabuľka S5. Jaccardove indexy meraní lokálnej centrality na porovnanie medzi PBS+ B. afzelii vs. E. coli+B. afzelii skupiny. Jaccardove indexy pre každú z mier lokálnej centrálnosti (tj stupeň, centrálnosť medziľahlosti, centrálnosť blízkosti, centrálnosť vlastného vektora a uzlové taxóny) súborov väčšiny centrálnych uzlov na porovnávanie párových sietí. Dve p-hodnoty, P(J ≤ j) a P(J ≥ j), pre každý Jaccardov index sa pridali
40168_2023_1599_MOESM13_ESM.xlsx (16,7 kB, xlsx)Doplnkový súbor 13: Doplnková tabuľka S6. Spoločné a jedinečné susedné uzly taxónu Escherichia-Shigella medzi PBS+ B. afzelii a E. coli+B. afzelii skupiny.
40168_2023_1599_MOESM14_ESM.csv (1,8 MB, csv)Doplnkový súbor 14: Doplnková tabuľka S7. Bakteriálne taxóny nachádzajúce sa ako kontaminanty v súboroch údajov o sekvenovaní génu 16S rRNA z kliešťov vystavených B. afzelii alebo E. coli+B. afzelii kapilárnym kŕmením alebo mikroinjekciou do análnych pórov. Kontaminanty boli štatisticky identifikované (TRUE) a odstránené z dátových súborov sekvenovania génu 16S rRNA pomocou balíka decontam R.
Vyhlásenie o dostupnosti údajov
Súbory údajov prezentované v tejto štúdii možno nájsť v online úložiskách. Názvy repozitárov/úložísk a prístupové čísla (čísla) nájdete nižšie: https://www.ncbi.nlm.nih.gov/sra , PRJNA870490.




