


RECENZNÝ článok
Patobiológia proteínu proti starnutiu Klotho a terapeutické úvahy


- 1 Oddelenie laboratórnej medicíny a patobiológie, University of Toronto, Toronto, ON, Kanada
- 2 Department of Laboratory Medicine, Keenan Research Center for Biomedical Science, Unity Health Toronto, Toronto, ON, Kanada
- 3 Oddelenie endokrinológie a metabolizmu, Nemocnica Huashan, Shanghai Medical School, Univerzita Fudan, Šanghaj, Čína
- 4 Shanghai Yinuo Pharmaceutical Co., Ltd., Šanghaj, Čína
Proteín a-Klotho (ďalej označovaný ako Klotho) má vlastnosti proti starnutiu, ako sa prvýkrát pozorovalo u myší homozygotných pre hypomorfný gén Klotho ( kl/kl ). Tieto myši majú skrátenú dĺžku života, zakrpatený rast, ochorenie obličiek, hyperfosfatémiu, hyperkalcémiu, vaskulárnu kalcifikáciu, srdcovú hypertrofiu, hypertenziu, pľúcne ochorenie, kognitívnu poruchu, multiorgánovú atrofiu a fibrózu. Nadmerná expresia Klotho má opačné účinky, predlžuje životnosť. U ľudí hladiny Klotho klesajú s vekom, chronickým ochorením obličiek, cukrovkou, Alzheimerovou chorobou a inými stavmi. Nízke hladiny Klotho korelujú so zvýšením úmrtnosti zo všetkých príčin. Klotho pôsobí buď ako obligátny koreceptor pre fibroblastový rastový faktor 23 (FGF23), alebo ako rozpustný pleiotropný endokrinný hormón (s-Klotho). Produkuje sa najmä v obličkách, ale aj v mozgu, pankrease a iných tkanivách. Na membránach tubulárnych buniek obličiek sa spája s receptormi FGF, aby naviazali FGF23. FGF23, produkovaný v kostiach, reguluje renálne vylučovanie fosfátu (fosfaturický účinok) a metabolizmus vitamínu D. Nedostatok Klotho alebo FGF23 vedie k hyperfosfatémii a hypervitaminóze D. S pribúdajúcim vekom sa funkcia obličiek u ľudí často zhoršuje, čím sa znižujú hladiny Klotho. Zdá sa, že to podporuje patológiu súvisiacu s vekom. Je pozoruhodné, že Klotho inhibuje štyri dráhy, ktoré sú spojené so starnutím rôznymi spôsobmi: transformujúci rastový faktor β (TGF-β), inzulínu podobný rastový faktor 1 (IGF-1), Wnt a NF-κB. Tieto môžu vyvolať starnutie buniek, apoptózu, zápal, imunitnú dysfunkciu, fibrózu a neopláziu. Okrem toho Klotho zvyšuje bunkové ochranné antioxidačné enzýmy prostredníctvom Nrf2 a FoxO. V súlade s tým predklinická liečba Klotho zlepšila obličkové, kardiovaskulárne, s cukrovkou a neurodegeneratívne ochorenia, ako aj rakovinu. Proteínová injekcia s-Klotho bola účinná, ale vyžaduje si ďalšie skúmanie. Niekoľko liekov zvyšuje hladiny cirkulujúceho Klotho a niektoré prechádzajú hematoencefalickou bariérou, aby potenciálne pôsobili v mozgu. V klinických štúdiách sa zvýšenie Klotho zaznamenalo pri inhibítoroch renín-angiotenzínového systému (losartan, valsartan), statíne (fluvastatín), inhibítoroch mTOR (rapamycín, everolimus), vitamíne D a pentoxifylíne. V predklinickej práci posilnili Klotho aj antidiabetiká (metformín, na báze GLP-1, GABA, PPAR-γ agonisty). Niekoľko tradičných liekov a/alebo nutraceutík zvýšilo Klotho u hlodavcov, vrátane astaxantínu, kurkumínu, ženšenu, ligustilidu a resveratrolu. Najmä cvičenie a športová aktivita zvýšili Klotho. Tento prehľad sa zaoberá molekulárnymi, fyziologickými a terapeutickými aspektmi Klotho.
1 Úvod
Starnutie je spojené so zmenami v takmer všetkých tkanivách a orgánoch tela, čo nakoniec vedie k oslabujúcim a/alebo smrteľným stavom, ako sú kardiovaskulárne ochorenia, chronické zlyhanie orgánov, neurodegenerácia a rakovina. Starnutie prebieha u rôznych druhov rôznou rýchlosťou, čo naznačuje niektoré zle definované biologické hodiny. Molekulárne mechanizmy starnutia sú intenzívne študované, ale stále nie sú dobre pochopené. Patogenéza chorôb spojených s vekom je však pravdepodobne multifaktoriálna ( Franceschi et al., 2018 ). U ľudí bol obmedzený počet génov jasne spojený so zrýchleným starnutím, ako je vidieť v progérii (zriedkavý stav) ( Coppede 2021 ) alebo naopak, v súvislosti s dlhovekosťou ( Javidnia et al., 2022 ). Predmetom tohto prehľadu je Klotho (kl) , čo je gén proti starnutiu, a zodpovedajúci proteín α-Klotho (ďalej označovaný ako Klotho alebo KL). Gén bol prvýkrát identifikovaný u myší v roku 1997 ( Kuro-o et al., 1997 ). Nedostatok proteínu vedie k syndrómu, ktorý má niekoľko znakov starnutia, ako bolo pozorované u mutantných myší buď s hypomorfnou Klotho alelou ( Kl kl/kl ) ( Kuro-o, et al., 1997 ) alebo úplným vyradením génu Klotho. ( Kl -/- ) ( Tsujikawa et al., 2003 ). Klotho-deficientné myši vykazujú zakrpatený rast, ochorenie obličiek, hyperfosfatémiu, hyperkalcémiu, vaskulárnu kalcifikáciu, srdcovú hypertrofiu, hypertenziu, fibrózu orgánov, multiorgánovú atrofiu, osteopéniu, pľúcne ochorenie, kognitívne poruchy a krátku životnosť ( Bian et al., 2015 ; Erben a Andrukhova, 2017 , Kinoshita a Kawai, 2016 , Kuro-o a kol., 1997 , 2010 , 2019 , 2021 ). Nadmerná expresia génu má opačné účinky, predlžuje prežitie ( Kurosu et al., 2005 ).
Zdá sa, že klotho insuficiencia hrá úlohu pri starnutí človeka a konkrétne pri mnohých chorobách, ktoré sú spojené so starnutím. Expresia Klotho klesá s vekom, zlyhaním obličiek, cukrovkou a neurodegeneratívnym ochorením. Zdá sa, že pokles hladín v sére súvisiaci s vekom je podobný u mužov a žien; a referenčné hodnoty boli nedávno publikované ( Espuch-Oliver et al., 2022 ). Nedávna štúdia dospelých Američanov ukázala, že nízke hladiny Klotho v sére korelujú so zvýšenou mierou úmrtnosti zo všetkých príčin ( Kresovich a Bulka, 2021 ).
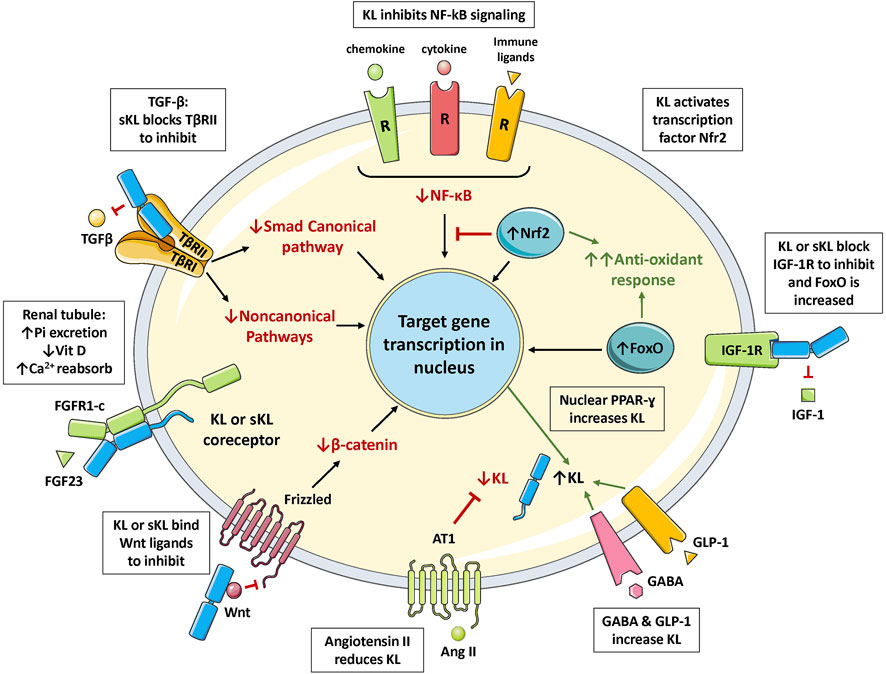
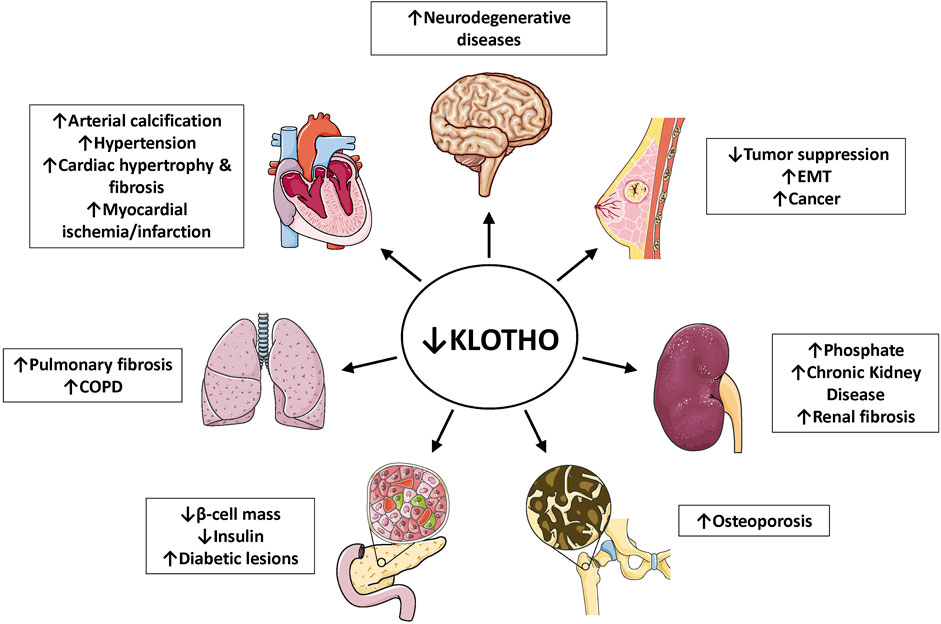
Klotho môže existovať ako membránovo viazaný coreceptor pre fibroblastový rastový faktor 23 (FGF23) ( Urakawa et al., 2006 ) alebo rozpustný endokrinný mediátor s mnohými funkciami ( obrázky 1 , 2 ) ( Dalton et al., 2017 ; Kuro- o, 2017 ). Zhoršenie funkcie obličiek súvisiace s vekom má za následok Klotho insuficienciu a hyperfosfatémiu, ktorá výrazne prispieva k fenotypu starnutia ( Kuro-o 2010 , 2018 , 2021 ). Klotho chráni obličky a podporuje vylučovanie fosfátov (fosfaturický efekt). Je pozoruhodné, že nezávisle od FGF23 inhibuje najmenej štyri dráhy, ktoré sú rôznymi spôsobmi spojené so starnutím. Klotho blokuje alebo inhibuje transformujúci rastový faktor p (TGF-p), inzulínu podobný rastový faktor 1 (IGF-1), jadrový faktor KB (NF-KB) a Wnt/p-katenín ( obrázok 3 ). V dôsledku toho, ako bude uvedené v tomto prehľade, Klotho má hlavné účinky na niekoľko biologických procesov súvisiacich so starnutím a chorobami ( obrázok 4 ):
1) Regulácia fosfátu, vápnika a vitamínu D závislá od FGF23.
2) Antioxidačné a protizápalové aktivity.
3) Prevencia chronickej fibrózy.
4) Ochranné účinky proti kardiovaskulárnym ochoreniam.
5) Protirakovinové (nádorové supresorové) aktivity.
6) Metabolické regulačné funkcie relevantné pre diabetes.
7) antiapoptotické a anti-senescenčné funkcie; zachovanie kmeňových buniek.
8) Ochrana pred neurodegeneratívnym ochorením (Alzheimer a iné).
Okrem týchto tém budeme informovať o niekoľkých faktoroch, ktoré zvyšujú Klotho, vrátane bežne predpisovaných liekov, rekombinantných proteínov, génových terapií, tradičných liekov, nutraceutík a cvičenia.
2 stránky výroby Klotho
Klotho sa primárne produkuje v obličkách (renálne tubuly), ale nachádza sa aj v mozgu (choroidný plexus, CSF a neuróny), pankreatických β bunkách, krvných cievach a koži (Lim K, et al., 2015). Nedávna práca tiež dokumentuje expresiu v periférnych krvných cirkulujúcich bunkách ( Martin-Nunez et al., 2022 ). Vylučuje sa aj do moču. Štúdie na nefrektomizovaných hlodavcoch ( Hu a kol., 2016 ) a myšiach s genetickou deléciou Klotho špecifickou pre obličky ( Lindberg a kol., 2014 ) odhalili, že obličky sú hlavným zdrojom cirkulujúceho Klotho. Produkujú ho proximálne a distálne stočené obličkové tubuly, možno viac v distálnych tubuloch (Lim K, et al., 2015). Niektorí autori však uvádzajú podobné množstvá membránovo viazanej formy na oboch miestach ( Andrukhova et al., 2012 ; Erben a Andrukhova, 2017 ). Je zaujímavé, že Klotho má rôzne funkcie v proximálnych a distálnych tubuloch ( Erben a Andrukhova, 2017 ). V proximálnom tubule Klotho podporuje fosfaturický účinok a inhibuje produkciu vitamínu D, zatiaľ čo v distálnom tubule zvyšuje reabsorpciu Ca2 + . Podmienený knockout génu Klotho v proximálnom tubule reprodukoval mnohé z vlastností systémového génového knockoutu, čo potvrdzuje dôležitosť proximálnych tubulárnych funkcií ( Takeshita et al., 2017 ).
3 Molekulárne vlastnosti a regulácia
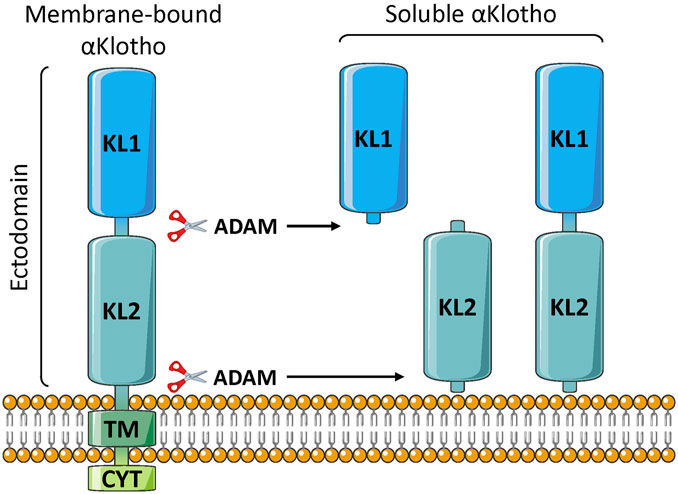
3.1 Membránovo viazané a rozpustné formy
Klotho pozostáva z jednopriechodového membránového proteínu 1 012 aa (130 kDa), u ľudí, s veľmi krátkym intracytoplazmatickým segmentom (10 aa) ( Xu a Sun 2015 ; Chen G, et al., 2018). Extracelulárna zložka pozostáva z dvoch domén, KL1 a KL2 ( obrázok 1 ). Extracelulárna časť môže byť štiepená membránovými proteázami, predovšetkým ADAM10 a ADAM17 (α-sekretázy), a táto rozpustná forma α-Klotho (s-Klotho) sa uvoľňuje do telesných tekutín, kde pôsobí ako endokrinný hormón ( Xu a Sun, 2015 Dalton a kol., 2017 ). Dodatočné štiepenie môže uvoľniť menšie fragmenty KL1 alebo KL2, hoci podľa našich vedomostí neboli v obehu detegované. Možno existuje aj iná rozpustná forma Klotho; o ktorých sa predpokladá, že sú generované alternatívnym zostrihom, pozostávajúcim iba z KL1 a často označovaných ako vylučovaný Klotho. Sekvencia však obsahuje predčasné stop kodóny a mRNA je degradovaná ( Mencke, et al., 2017a ). Zdá sa teda, že prevažná väčšina alebo možno všetky rozpustné Klotho pozostávajú z formy KL1/KL2 odvodenej proteolytickým štiepením proteínu viazaného na membránu (vylučovanie). V skutočnosti u hlodavcov podávanie inhibítorov sekretáz sprostredkujúcich toto proteolytické štiepenie drasticky znižuje hladiny s-Klotho v obehu ( Hu et al., 2016 ).
Regulácia vylučovania Klotho nie je úplne objasnená, ale úloha a-sekretázy ADAM10 a ADAM17 je dobre zdokumentovaná, najmä v renálnych tubulárnych epiteliálnych bunkách ( Van Loon et al., 2015 ). Úroveň expresie týchto enzýmov môže byť zvýšená rôznymi spôsobmi, napríklad pôsobením inzulínu, rastových faktorov a cytokínov ( Bzowska a kol., 2004 ; Chen CD, a kol., 2007 ; Maretzky a kol., 2008 ; Murphy , 2009 ). Naproti tomu tkanivové inhibítory metaloproteináz (TIMP) blokujú pôsobenie ADAM proteínov ( Chen CD, et al., 2007 ; Murphy, 2009 ). Niektoré lieky môžu zvýšiť proteolytické štiepenie Klotho. Publikovaných príkladov je málo; pozoruhodným prípadom je však ligustilid. Táto prírodná zlúčenina zvýšila expresiu ADAM10 a s-Klotho a bola ochranná na myšom modeli Alzheimerovej choroby (pozri časť 10.4) ( Kuang et al., 2014 ). Je dôležité poznamenať, že sekretázové proteíny ADAM majú veľmi veľký počet substrátov a môžu modifikovať mnohé biologické procesy. Preto liekom zameraným na tieto enzýmy pravdepodobne chýba špecifickosť a podliehajú toxicite.
Bola publikovaná kryštálová štruktúra Klotho a jeho väzbové miesta k receptorom FGF (najmä FGFR1c) a FGF23 (Chen G, et al., 2018). Nebol identifikovaný žiadny špecifický receptor pre s-Klotho. Viaže sa však na niektoré receptory, ktoré sú široko distribuované na mnohých typoch buniek; najmä FGFR a receptor TGF-p.
3.2 Regulácia expresie Klotho
Niekoľko faktorov reguluje expresiu Klotho, ako sa uvádza v recenzii ( Xu a Sun, 2015 ). To sa môže v rôznych tkanivách značne líšiť a je to len čiastočne pochopené. V obličkách, ktoré sú hlavným miestom produkcie, môžu rôzne faktory meniť expresiu Klotho za fyziologických a patologických podmienok, ako je obehový stres, hypertenzia, oxidačný stres a diabetes. Klotho je vo všeobecnosti depresívny pri zápalových poruchách a je potenciálne užitočným biomarkerom zápalu ( Wu a Chen, 2022 ).
Viaceré transkripčné faktory regulujú expresiu génu Klotho , vrátane tých, ktoré sa viažu na promótorové miesta E-box, Ap-2, PAX4, Sp1 a Oct-1 ( Xu a Sun, 2015 ). PAX4 je pozitívny regulátor. V 5′ ohraničujúcej oblasti génu sú prvky odozvy PPAR-y, ktoré zvyšujú transkripciu. PAX4 a PPAR-y sú relevantné pre diabetes, ako je uvedené v iných častiach. Vitamín D sa viaže na prvky odozvy vitamínu D v promótore, aby upreguloval transkripciu Klotho . Na rozdiel od toho, NF-KB sa viaže na promótor Klotho , ale je inhibičný ( Moreno et al., 2011 ; Lin et al., 2021 ). To by mohlo aspoň čiastočne vysvetliť znížené hladiny Klotho pri zápalových stavoch. Najmä angiotenzín II je hlavným inhibítorom transkripcie Klotho ( obrázok 3 ), čo sa pripisuje supresii pozitívneho regulátora Sp1 ( Zhou Q, et al., 2010 ). HMG-CoA reduktáza je tiež inhibičná a to sa dá zvrátiť statínovými liekmi, ktoré sú inhibítormi tohto enzýmu ( Narumiya et al., 2004 ; Kuwahara et al., 2008 ) ( tabuľka 2 ). Statíny upregulujú Klotho mRNA, pravdepodobne inaktiváciou RhoA a/alebo znížením odpovede angiotenzínu II ( Narumiya et al., 2004 ). Je zaujímavé, že FGF23 je silný negatívny regulátor transkripcie Klotho ( Marsell et al., 2008 ). V dôsledku toho udalosti, ktoré zvyšujú produkciu FGF23, znížia Klotho, ako je chronické ochorenie obličiek ( Kuro-o, 2019 ). Gén Klotho je v rakovinových bunkách často vypnutý, možno prostredníctvom množstva mechanizmov ( Rubinek a Wolf, 2016 ). Najmä promótor Klotho je bohatý na GC a podlieha metylácii DNA a v nádoroch môže byť vypnutý hypermetyláciou DNA. Klotho je tiež regulované miRNA a zdá sa, že to je dôležité pri rakovine ( Abolghasemi et al., 2019 ).
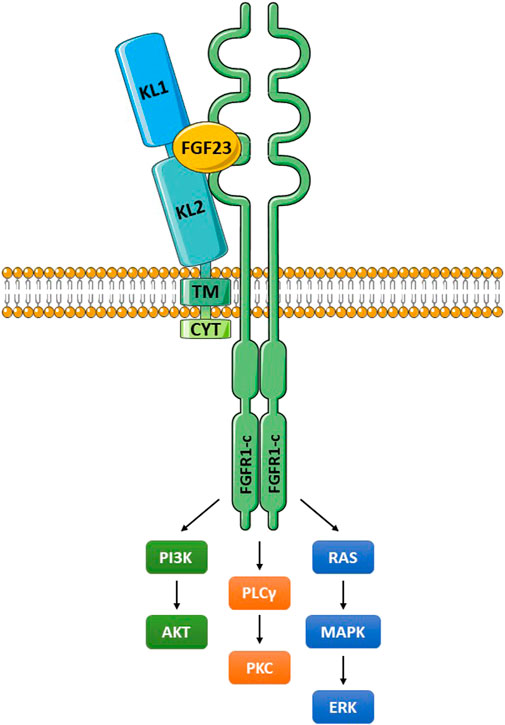
4 Klotho interakcie s FGF23 a FGFR
Extracelulárna doména Klotho sa viaže buď na FGFR1c, -3c alebo -4 (všetky receptorové tyrozínkinázy), čím sa vytvorí vysoko afinitný receptor pre FGF23 ( obrázok 2 ) (Chen G, et al., 2018; Urakawa et al., 2006 ). Pokiaľ ide o nomenklatúru, FGF19, FGF21 a FGF23 sú často označované endokrinné FGF (eFGF), pretože sú rozpustné, vstupujú do obehu a môžu pôsobiť na diaľku ( Phan et al., 2021 ). FGF23 je eFGF produkovaný v kostiach, ktorý reguluje renálnu fosfátovú homeostázu spolu s vitamínom D a parathormónom (PTH) ( Neyra et al., 2021 ; Rausch a Föller, 2022 ). Membránovo viazané a rozpustné formy Klotho môžu fungovať ako koreceptory pre FGF23 (Chen G, et al., 2018). Bez Klotho je afinita FGF23 k receptoru veľmi nízka. Aktivovaný FGFR signalizuje cez PI3K/Akt, fosfolipázu Cy (PLCy) a Ras/MAPK/ERK. Renálne fyziologické funkcie Klotho boli podrobne preskúmané ( Erben a Andrukhova, 2017 ; Kuro-o 2017 ; Zou a kol., 2018 ; Kuro-o 2019 ; Kuro-o 2021 ; Neyra a kol., 2021 ; Saar-Kov al., 2021 ) a sú tu opísané len stručne. Aktivácia FGFR1c reguluje výmenu fosfátov a vápnika v obličkách. K tomu dochádza prostredníctvom inhibície transportérov sodíka a fosforečnanu NPT-2a a NPT-2c v proximálnom renálnom tubule, čím sa znižuje reabsorpcia anorganického fosfátu (Pi) (fosfaturický efekt) ( Erben, 2018 ). V distálnom stočenom tubule je zvýšená resorpcia Ca2 + cez TRPV5 kanály. V distálnom tubule je tiež zvýšená reabsorpcia Na + prostredníctvom zvýšenej expresie Na + :Cl − kotransportéra (NCC). Hlavný fyziologický význam má FGF23/Klotho/FGFR1c , ktorý potláča expresiu la-hydroxylázy v proximálnom renálnom tubule, čo inhibuje syntézu aktívneho vitamínu D, označovaného ako 1,25(OH) 2D3 ( kalcitriol). Okrem iných účinkov vitamín D podporuje vstrebávanie fosfátov a vápnika v čreve, a to sa zníži. Okrem toho, spôsobom nezávislým od Klotho, FGF23 znižuje sekréciu parathormónu (PTH), ktorý navyše ovplyvňuje rovnováhu fosforečnanu a vápnika ( Ben-Dov et al., 2007 ).
Nedostatok Klotho alebo FGF23 u myší má za následok nadmernú aktivitu 1α-hydroxylázy, nadprodukciu aktívneho vitamínu D a súvisiacu hyperfosfatémiu a hyperkalcémiu. U myší s deficitom Klotho sa zdá, že hypervitaminóza D a hyperfosfatémia prispievajú k fenotypu zrýchleného starnutia ( Kuro-o 2010 , 2018 , 2019 , 2021 ). Avšak diéta s nízkym obsahom fosfátov zlepšuje ochorenie, aj keď vitamín D ďalej stúpa, čo naznačuje, že hyperfosfatémia je pri syndróme starnutia dôležitejšia ako toxicita vitamínu D. Napriek tomu ablácia odpovedí na vitamín D zoslabila lézie myší s deficitom Klotho ( Anor et al., 2012 ). Bolo navrhnuté, že hypersaturácia fosfátových a vápenatých iónov v krvi vedie k tvorbe kalciproteínových častíc (CPP) ( Kuro-o, 2021 ). CPP sa zrážajú v tkanivách a predpokladá sa, že urýchľujú patologické zmeny spojené so starnutím.
Najčastejšou klinickou príčinou nedostatku Klotho je zlyhanie obličiek, ktoré môže byť výsledkom mnohých akútnych alebo chronických stavov ( Neyra et al., 2021 ). Zriedkavé ľudské mutácie spôsobujúce závažný nedostatok buď Klotho alebo FGF23 sú spojené s hyperfosfatémiou začínajúcou v ranom veku ( Ito a Fukumoto, 2021 ). Ide o autozomálne recesívne ochorenia. U subjektov sa rozvinie masívna kalcifikácia v mäkkých tkanivách, krvných cievach a vo viacerých orgánoch a anatomických miestach v celom tele. Existujú kostné a zubné lézie. Existujú aj dôkazy o systémovom zápale. Liečba zahŕňa diétu s nízkym obsahom fosfátov, fosfaturové lieky, protizápalové lieky a látky viažuce fosfáty. Tento závažný vrodený nedostatok funkčného Klotho alebo FGF23 nie je porovnateľný s oveľa menej zníženou expresiou Klotho pozorovanou pri chronických ochoreniach opísaných v tomto prehľade. V skutočnosti nie sú spojené s masívnou kalcifikáciou pozorovanou pri vrodenej hyperfosfatémii.
5 β-Klotho
Homológ Klotho označený ako β-Klotho (KLB) je povinným koreceptorom pre FGF19 a FGF21, čo sú oba eFGF ( Kilkenny a Rocheleau., 2016 ). Zdá sa, že KLB, na rozdiel od Klotho, nemá endokrinné funkcie ako také. FGF19 hrá hlavnú úlohu v homeostáze žlčových kyselín a metabolizme pečene ( Dolegowska et al., 2019 ; Guthrie et al., 2022 ). FGF21 sa podieľa na regulácii metabolizmu lipidov, sekrécie inzulínu a homeostázy glukózy ( Andersen a kol., 2015 ; Bondurant a Potthoff, 2018 ; Dolegowska a kol., 2019 ). FGF21 sa tvorí v pečeni, pankreatických ostrovčekoch, exokrinnom pankrease a iných tkanivách. Zdá sa, že pečeň je zodpovedná za väčšinu cirkulujúceho FGF21. Hlavným metabolickým funkciám FGF21 bola venovaná značná pozornosť a bol skúmaný v klinických skúškach na liečbu obezity a cukrovky.
6 Klotho funkcií nezávislých od FGF23
6.1 Klotho inhibuje TGF-p
Kľúčovou ochrannou funkciou Klotho je prevencia fibrózy (napr. pľúcnej, renálnej alebo srdcovej fibrózy), ktorá sa z veľkej časti pripisuje jeho schopnosti blokovať TGF-β, hoci prispievajú aj iné cesty ( Huang Q, et al., 2020 , Mencke a kol., 2017b). TGF-β je pleiotropný cytokín, ktorý je spojený so starnutím jeho podporou starnutia buniek, úbytkom kmeňových buniek, imunologickým poškodením, fibrózou a niekoľkými ďalšími patológiami súvisiacimi s vekom ( Mikuła-Pietrasik et al., 2022 ; Pratsinis et al., 2017 ; Prud’homme 2007 , a kol., 2017b ; a Suzuki, 2019 ). Toto je obzvlášť zaujímavé, pretože Klotho sa viaže na receptor TGF-β typu II (TβRII), ktorý blokuje väzbu TGF-β a signalizáciu receptora ( Doi et al., 2011 ) ( obrázok 3 ). V obličkách starnúcich myší hladiny Klotho klesajú, zatiaľ čo TGF-β a jeho signálne molekuly sa zvyšujú ( Oishi et al., 2021 ). Podávanie proteínu s-Klotho inhibuje signalizáciu TGF-β a chráni pred renálnou fibrózou ( Doi et al., 2011 ). Špecifickejším spôsobom, peptid odvodený od Klotho (30 aminokyselín), ktorý inhibuje TGF-p väzbou na jeho receptor, tiež chráni pred renálnou fibrózou ( Yuan et al., 2022 ). Tieto experimenty poskytujú dôkaz, že Klotho môže potlačiť aktivity TGF-p in vivo .
Prakticky všetky typy buniek exprimujú receptory TGF-β a jeho význam je značný. Signalizácia TGF-β a jej regulácia sú zložité ( Mullen a Wrana, 2017 ; Derynck a Budi, 2019 ; Aashaq a kol., 2022 ) a je tu uvedený len stručný prehľad. TGF-p je vylučovaný v latentnej forme a je aktivovaný interakciou s integrínmi a inými mechanizmami. Existujú tri izoformy TGF-β (TGF-β1 je najhojnejší), ktoré sa všetky viažu na rovnaký signálny receptor ( Groppe a kol., 2008 ; Huang T, a kol., 2011 ; Aashaq a kol., 2022 ). Tento receptor pozostáva z TβRI (tiež označovaného ALK5) a TβRII. TGF-β sa viaže na TβRII a TβRI za vzniku komplexu serín/treonín kinázy. TβRII fosforyluje TβRI, ktorý potom fosforyluje Smad2 a Smad3 (kanonická dráha). Smad2 a 3 tvoria komplex so Smad4 (bežný Smad), ktorý sa translokuje do jadra. Tam sa viaže na DNA a reguluje transkripčné udalosti. Viaceré molekuly interagujú s komplexom receptora TGF-p a môžu meniť väzbu TGF-p alebo receptorovú signalizáciu pozitívnym alebo negatívnym spôsobom ( Pawlak a Blobe, 2022 ; Prud’homme et al., 2017b ). Napríklad buď betaglykán (tiež označovaný ako TβRIII), neuropilín-1 (Nrp1) alebo endoglín sa môžu spájať s receptorom TGF-β, pôsobiacim ako koreceptory a charakteristicky zosilňujú reakcie TGF-β. Klotho sa tiež viaže na tento receptor, ale má opačný účinok.
Aktivácia niekoľkých nekanonických (nezávislých od Smad2/3) dráh pridáva väčšiu komplexnosť ( Derynck a Budi, 2019 ; Aashaq a kol., 2022 ; Baba a kol., 2022 ). V niektorých bunkách, najmä v endoteliálnych bunkách, môže k signalizácii dôjsť prostredníctvom alternatívnej dráhy ALK1/Smad1,5,8. Smad4 (bežný Smad) sa potom zúčastňuje rovnako ako v ceste Smad2/3. Signalizačný TGF-p receptor môže aktivovať ERK, JNK, p38 MAPK, PI3K/Akt, Rho-podobné GTPázy, NF-KB a ďalšie dráhy. Okrem toho, prostredníctvom kanonickej alebo nekanonickej signalizácie môže TGF-P prepočúvať s kostnými morfogenetickými proteínmi (BMP), Wnt, Hedgehog, Notch, Hippo (TAZ/YAP), JAK/STAT; ako aj rastové faktory, ako je hepatocytový rastový faktor (HGF) a epidermálny rastový faktor (EGF) ( Luo 2017 ; Mullen a Wrana, 2017 ; Derynck a Budi, 2019 ). TGF-β teda môže mať vplyv na veľké množstvo biologických procesov. V imunitnom systéme ho produkujú regulačné T bunky (Treg) a iné typy buniek a potláča alebo reguluje imunitné reakcie zahŕňajúce makrofágy, dendritické bunky, B bunky, efektorové T bunky, NK bunky a neutrofily ( de Streel a Lucas, 2021 ; Je teda kľúčovým regulátorom imunitného systému.
Štúdie Klotho inhibície TGF-p uvedené vyššie sa týkajú primárne ochorenia obličiek, ale sú pravdepodobne použiteľné aj na fibrózu inde, ako je pľúcna fibróza a niekoľko ďalších stavov súvisiacich s vekom. To by bolo v súlade so všadeprítomnou expresiou a viacerými funkciami TGF-p a jeho receptorov. TGF-p sa podieľa na mnohých léziách pozorovaných pri cukrovke. Úloha TGF-β pri neoplázii je komplexná a závisí od kontextu. V skorých štádiách neoplázie pôsobí ako nádorový supresor, ale zhoršuje rakovinu v neskorých fázach a prispieva k metastatickému ochoreniu ( de Streel a Lucas 2021 ; Baba et al., 2022 ). Preto je jeho inhibícia Klotho relevantná pre liečbu rakoviny.
6.2 Klotho Inhibuje NF-KB
Predpokladá sa, že zápal je hlavným prispievateľom k starnutiu, v procese niekedy označovanom ako zápal ( de Almeida a kol., 2020 ; Fulop a kol., 2021 ). Chronický zápal nízkeho stupňa môže viesť k trvalému poškodeniu tkaniva. Napríklad zápalové zmeny rôznych typov sú spojené s aterosklerózou, chronickým ochorením obličiek, poškodením orgánov súvisiacim s cukrovkou a Alzheimerovou chorobou. Klotho vykazuje protizápalové aktivity, ktoré sa zdajú byť nezávislé od FGF23. Dôležité je, že Klotho potláča aktiváciu zápalovej dráhy NF-KB ( Buendia et al., 2015 , 2016 ). Táto dráha hrá kľúčovú úlohu pri iniciácii imunitných a/alebo zápalových reakcií sprostredkovaných B bunkami, T bunkami, makrofágmi a polymorfonukleárnymi leukocytmi (PML) ( Haga a Okada, 2022 ; Roberti et al., 2022 ). Inhibuje tiež apoptózu imunitných buniek a iných typov buniek, pričom podporuje proliferáciu. Je aktivovaný kostimulačnými receptormi T-buniek a B-buniek, niekoľkými zápalovými cytokínmi, chemokínmi, toll-like receptormi (TLR), NOD-like receptormi (NLR), stimulátorom interferónových génov (STING) a ďalšími faktormi, ktoré podporujú imunitu proti infekčným agens ( Roberti a kol., 2022 ; Haga a Okada, 2022 ; Zhang T, a kol., 2021). NF-KB sa však podieľa aj na škodlivom zápale, ktorý prispieva k starnutiu, chronickým zápalovým stavom a autoimunitným ochoreniam. NF-KB je aktívny v mnohých neimunitných typoch buniek, ako sú endotelové bunky a niektoré epitelové bunky. Prispieva k vaskulárnym léziám (napr. ateroskleróza a vaskulitída) a čo je dôležité, hrá škodlivú úlohu pri rakovine (Zhang T, et al., 2021). NF-KB teda môže prispievať k chorobám súvisiacim s vekom mnohými spôsobmi.
Rodina NF-KB zahŕňa NF-KB1 (p50), NF-KB2 (p52), RelA (p65) a RelB a nedávno boli preskúmané aktivačné dráhy NF-KB ( Roberti et al., 2022 ; Haga a Okada 2022 , Zhang T, a kol., 2021). Všetky tieto proteíny majú Rel homológnu doménu pre sekvenčne špecifickú väzbu DNA, ako aj homo- a heterodimerizáciu. Existujú dve cesty pre signalizáciu, označované ako kanonické a nekanoické (alternatívne) cesty. Včasné signalizačné udalosti v dráhach NF-KB zahŕňajú aktiváciu kináz, ako je TAK1 a komplex NEMO. V kanonickej dráhe, v kľudovom stave, je inhibítor κB (IκB) naviazaný na NF-κB (p50/RelA) a to bráni jeho aktivácii. V reakcii na stimulačný účinok je IκB proteín fosforylovaný, ubikvitinovaný a podrobený proteazomálnej proteolýze. To umožňuje NF-KB migrovať do jadra a viazať sa na DNA a indukovať transkripciu veľkého počtu génov. V nekanonickej dráhe sa RelB viaže na proteín označený p100. Aktivácia kinázy indukujúcej NF-KB (NIK) vedie k fosforylácii proteínu p100 a jeho následnej degradácii na proteín p52. Potom sa komplex p52/RelB translokuje do jadra a aktivuje transkripciu cieľových génov. Je zaujímavé, že existujú dôkazy, že tieto dve cesty sa vzájomne ovplyvňujú a nemusia byť úplne nezávislé.
Hlavné inhibičné funkcie Klotho vyplývajú z jeho schopnosti blokovať signalizáciu NF-KB, ako sa preukázalo v niekoľkých štúdiách. V tomto prípade sa zdá, že Klotho bráni jadrovej translokácii NF-KB, hoci sa môžu uplatniť aj iné mechanizmy, ako je opísané nižšie. Napríklad Klotho potlačil TNF-a-indukovanú aktiváciu NF-KB v endotelových bunkách ( Maekawa et al., 2009 ). Urémia je spojená s cirkulujúcimi toxínmi, ktoré spôsobujú oxidačný stres endotelovým bunkám a vyvolávajú ich starnutie. Tieto endotelové zmeny sa pripisovali aktivácii NF-KB, čo Klotho zmiernilo ( Buendia et al., 2015 , 2016 ). Autori dospeli k záveru, že Klotho zabraňuje jadrovej translokácii NF-κB, čo je nevyhnutný krok v aktivačnej dráhe. Iní vyšetrovatelia hlásili podobné zistenia ( Yang, et al., 2012 ). V modeli diabetickej kardiomyopatie Klotho inhiboval NF-KB in vitro a in vivo ( Guo et al., 2018 ). Zdá sa, že toto potlačenie je spôsobené zvýšením aktivácie faktora 2 súvisiaceho s jadrovým faktorom 2 (Nrf2), ktorý pôsobí proti NF-KB (pozri nižšie). U diabetických db/db myší bola expresia Klotho znížená v obličkách, čo je zistenie spojené so zvýšenou aktiváciou NF-KB ( Zhao Y, et al., 2011 ). To bolo zvrátené aplikáciou Klotho s následným znížením zápalových cytokínov. Zistilo sa tiež, že Klotho inhibuje aktiváciu a translokáciu NF-KB v klonálnych p bunkách ( Prud’homme et al., 2017a ). V tomto prípade Klotho knockdown so siRNA viedol k spontánnej jadrovej translokácii NF-KB p65. To je v súlade s Klotho, ktorý bráni degradácii proteínu IκB, ako navrhli iní, ktorí študujú alveolárne makrofágy (Li L, et al., 2015). Je zaujímavé, že s-Klotho pridaný do kultúr obracia účinky Klotho knockdownu a zabraňuje aktivácii NF-KB (Li L, et al., 2015; Prud’homme et al., 2017a ). Mechanizmy inhibície NF-KB v týchto rôznych štúdiách sa môžu líšiť a nie sú úplne objasnené ( Typiak a Piwkowska, 2021 ). Okrem toho, akonáhle je NF-KB aktivovaný a translokovaný do jadra, môže sa viazať na promótor Klotho a inhibovať expresiu, ako už bolo uvedené ( Moreno et al., 2011 ; Lin et al., 2021 ). Takže pri silných akútnych alebo chronických zápalových stavoch môže byť aktivácia NF-KB dominantná a v dôsledku toho môže byť expresia Klotho znížená.
6.3 Klotho Aktivuje antioxidačnú dráhu Nrf2
Klotho aktivoval Nrf2 v modeloch renálnych, kardiovaskulárnych a neurologických predklinických ochorení ( Maltese et al., 2017 ; Zhu et al., 2017 ; Xing et al., 2021 ; Xiang et al., 2022 ). Nrf2 je transkripčný faktor, ktorý riadi reakcie na oxidačný stres a toxíny ( Dinkova-Kostova et al., 2018 ; Qu et al., 2020 ; Panda et al., 2022 ). Jeho domény Neh2 interagujú s negatívnym regulátorom označeným Kelch-like ECH-asociated protein 1 (Keap1) ( Panda et al., 2022 ). Keap1 sa viaže na Nrf2, čím podporuje jeho ubikvitináciu a proteazomálnu degradáciu. Vplyvom oxidačného stresu (alebo iných aktivátorov) sú tiolové skupiny na cysteínových zvyškoch v Keap1 modifikované a jeho funkcia je negovaná, čo vedie k uvoľneniu Nrf2. Potom sa Nrf2 premiestni do jadra a vytvorí heterodiméry s inými proteínmi a viaže sa na sekvenciu zosilňovača nazývanú prvok antioxidačnej odozvy (ARE). ARE sa podieľa na expresii génov bunkovej obrany, ktoré kódujú napríklad niekoľko antioxidačných proteínov a koenzým konjugovaný s glutatiónom. Existuje aj alternatívna cesta regulácie Nrf2, ktorá nevyžaduje Keap1. Nrf2 chráni pred oxidačným poškodením a, čo je dôležité, inhibuje zápalovú dráhu NF-KB ( Gao W, et al., 2022 ). Aktivácia Nrf2 pomocou Klotho sa javí ako dôležitý faktor pri ochrane pred obličkovými, cievnymi a inými ochoreniami.
6.4 Inhibícia dráhy IGF-1
Signálna dráha inzulín/IGF-1 receptora je už dlho spojená so starnutím a je citlivá na živiny ( Mathew et al., 2017 ; Johnson 2018 ). Keď je dostatok živín, aktivuje sa následný mediátor označený ako mechanický cieľ rapamycínu (mTOR; proteínkináza) a podieľa sa na niekoľkých poruchách súvisiacich s vekom. Obmedzenie kalórií (CR) môže zmierniť tento účinok a predĺžiť život u niektorých druhov ( Kapahi a kol., 2017 ; von Frieling a Roeder, 2020 ). Je teda veľmi dôležité, že Klotho inhibuje dráhu IGF-1/PI3K/Akt/mTOR ( Kurosu et al., 2005 :; Xie et al., 2013 ). Toto je jeden pravdepodobný mechanizmus, ktorým Klotho uplatňuje svoj účinok proti starnutiu a ochranu proti degeneratívnym ochoreniam ( Fung et al., 2022 ). V tomto prípade s-Klotho blokuje aktiváciu inzulínu/IGF-1 receptora, čím bráni následným signálnym udalostiam vrátane fosforylácie substrátov inzulínového receptora (IRS) a signalizácie PI3K/Akt/mTOR. Liek inhibujúci mTOR rapamycín odďaľuje ochorenie súvisiace s vekom, ako je vaskulárna kalcifikácia u myší, a čo je zaujímavé, zvyšuje expresiu Klotho (Zhao Y, et al., 2015). Inzulín/IGF-1 dráha tiež hrá úlohu v progresii rakoviny a jej inhibícia Klotho je nádorová supresívna.
Inzulínová/IGF-1 dráha sa spája s antioxidačnými mechanizmami prostredníctvom transkripčných faktorov FoxO forkhead (FOXO). Blokáda inzulínových/IGF-1 dráh uvoľňuje inhibíciu FOXO, čo vedie k ich jadrovej migrácii a expresii niekoľkých génov kódujúcich antioxidačné enzýmy, ako je napríklad mangán superoxiddismutáza ( Yamamoto et al., 2005 ). V súlade s tým je aktivácia FOXO potlačená u myší s deficitom Klotho, ale zvýšená u myší s nadmernou expresiou Klotho.
6.5 Inhibícia dráhy Wnt
Wnt je ďalšou hlavnou dráhou blokovanou Klotho (Liu H, et al., 2007). Wnt je signalizačná kaskáda zapojená do embryogenézy, biológie kmeňových buniek, špecifikácie bunkového osudu, polarity, mitózy a migrácie, ako bolo nedávno preskúmané ( Hayat a kol., 2022 ; Rim a kol., 2022 ). Dysregulácia Wnt má za následok anomálie vývoja, rôzne degeneratívne stavy a rakovinu. Existuje kanonická dráha závislá od β-katenínu a nekanonická (nezávislá od β-katenínu) dráha ( Hayat et al., 2022 ). Nekanonická má dva podtypy, tj polaritu planárnej bunky a dráhy Wnt/Ca2 + . V kanonickej signalizácii sú viaceré Wnt ligandy schopné viazať sa na príbuzný receptor, označovaný ako Frizzled, aby iniciovali reakciu. V neprítomnosti Wnt ligandu je dráha neaktívna. Je to spôsobené neustálou fosforyláciou cytoplazmatického β-katenínu sprostredkovanou proteínovým komplexom, čo vedie k jeho eliminácii proteazomálnou degradáciou. Avšak po naviazaní Wnt ligandu na Frizzle a koreceptor LRP sa degradácia β-katenínu ukončí a môže migrovať do jadra, aby aktivoval cieľové gény prostredníctvom interakcií s väzbovým faktorom T-bunkového faktora/lymfoidného zosilňovača ( TCF/LEF) transkripčné faktory. Táto kanonická dráha reguluje hlavne bunkovú proliferáciu, vrátane cyklínu D1, c-myc a mnohých ďalších proteínov; zatiaľ čo nekanonické dráhy regulujú polaritu a pohyblivosť buniek ( Hayat et al., 2022 ). Proliferačné aspekty sú obzvlášť relevantné pre rakovinu, ako je rakovina hrubého čreva a hepatocelulárny karcinóm.
Klotho blokuje aktiváciu Wnt väzbou na niekoľko Wnt ligandov, vrátane Wnt1, Wnt3, Wnt4 a Wnt5a (Liu H, et al., 2007; Wang a Sun, 2009 ). U myší s deficitom Klotho nadmerná aktivácia Wnt podporuje starnutie buniek a má negatívny vplyv na prežitie kmeňových buniek ( Bian et al., 2015 ). V obličkách je nadmerná expresia Wnt spojená s fibrózou (Li X, et al., 2021) a v tomto procese spolupracuje s TGF-β. V tomto ohľade môže Wnt spolupracovať aj s TGF-β na vyvolanie epitelovo-mezenchymálneho prechodu (EMT), ktorý je prekurzorom fibrózy (Li X, et al., 2021). Okrem toho bola EMT tiež spojená s diferenciáciou rakovinových kmeňových buniek (CSC) ( Prud’homme, 2012 ; Prud’homme, 2017b ). CSC sú vysoko tumorigénne, podporujú metastázy a odolávajú chemoterapii.
7 Význam cukrovky a obezity
Zaujímavé je, že Klotho je vyčerpaný na ostrovčekoch diabetických pacientov ( Lin a Sun, 2015a ). Niekoľko výskumníkov hlásilo znížené hladiny cirkulujúceho Klotho u jedincov s diabetom 1. typu (T1D) ( Keles a kol., 2016 ; Tarhani a kol., 2020 ; Zubkiewicz-Kucharska, 2021 ); a diabetes typu 2 (T2D), najmä ak ide o pokročilé ochorenie ( Nie a kol., 2017 ; Fountoulakis a kol., 2018 ; Zhang a Liu, 2018 ). Cirkulujúce hladiny s-Klotho sú znížené u db/db myší (T2D) ( Takenaka et al., 2011 ) a diabetických NOD myší (T1D) ( Prud’homme et al., 2020 ). Existujú dôkazy, že Klotho hrá dôležitú úlohu v metabolizme glukózy a lipidov, ako sa uvádza v prehľade ( Razzaque, 2012 ; Donate-Correa a kol., 2016 ; Wan Q, a kol., 2017 ; Landry a kol., 2021a ). Klotho-deficientné myši vykazujú atrofiu ostrovčekov a zníženú produkciu inzulínu ( Razzaque, 2012 ). Klotho zvyšuje expresiu β-buniek TRPV2 a zvyšuje vstup Ca2 + a reakciu indukovanú glukózou ( Lin a Sun, 2012 ).
FGFR1c je exprimovaný p bunkami a jeho oslabenie vedie k cukrovke a zníženému počtu p buniek ( Hart et al., 2000 ). Klotho aj KLB sú exprimované ostrovčekovými bunkami, čo naznačuje, že tieto bunky môžu reagovať na FGF23 (Klotho coreceptor) a FGF21 (KLB coreceptor). FGFR1c podporuje signalizáciu prostredníctvom niekoľkých kľúčových dráh pre P bunky ( obrázok 2 ), napr. PI3K/Akt (podporuje prežitie buniek), RAS-MAPK (podporuje mitogénnu odpoveď), PLC-PPAR-adiponektín (reguluje metabolizmus glukózy a lipidov). V súlade s tým Klotho podávaný génovým prenosom u myší preukázal hlavné ochranné účinky na β bunky v modeloch T1D alebo T2D ( Lin a Sun, 2015a , b ). Dôležité je, že Klotho znížil apoptózu β-buniek a zvýšil proliferáciu týchto buniek. Kľúčovým patologickým nálezom u NOD myší je rozvoj inzulitídy. Toto sa najprv objaví ako periostrovčekový infiltrát mononukleárnych buniek (vrátane T buniek), po ktorom nasleduje fokálna invázia imunitných buniek do ostrovčekov a následne závažná infiltrácia a strata β-buniek. Prenos génu Klotho redukoval inzulitídu, čo naznačuje imunosupresívny a/alebo protizápalový účinok. Okrem toho rekombinantný Klotho zlepšil ochorenie obličiek a hypertenziu u db/db myší ( Takenaka et al., 2019 ). Autori dospeli k záveru, že Klotho inhiboval TGF-β a signalizáciu tumor nekrotizujúceho faktora (TNF) na zníženie renálnej fibrózy.
Systémová liečba kyselinou y-aminomaslovou (GABA) chráni pred cukrovkou 1. typu (T1D) u myší ( Tian a kol., 2004 ; Soltani a kol., 2011 ; Wan Y, a kol., 2015 ; Wang a kol., 2019 ) . Indukuje replikáciu/regeneráciu myších a ľudských β-buniek, pričom znižuje apoptózu týchto buniek (Liu W, et al., 2017, 2021; Purwana et al., 2014 ; Sarnobat et al., 2022 ; Soltani et al., 2011 ; Tian a kol., 2013 ; al., 2019 ). Pokiaľ ide o mechanizmy, zistili sme, že GABA zvyšuje produkciu Klotho v modeli T1D s viacerými nízkymi dávkami streptozotocínu (STZ) ( Prud’homme et al., 2017a ). Liečba GABA zvýšila Klotho v pankreatických β bunkách, obličkách a plazme. Podobne v prípade ľudských ostrovčekov transplantovaných do imunodeficientných myší sme pozorovali, že perorálna liečba GABA zvýšila expresiu a replikáciu β-buniek Klotho a znížila apoptózu (Liu W, et al., 2021). Tieto účinky sa zlepšili spoločným podávaním inhibítora dipeptidylpeptidázy 4 (DPP-4).
U autoimunitných myší NOD náchylných na diabetes zvýšila injekcia proteínu s-Klotho pankreatickú β-bunkovú replikáciu a β-bunkovú hmotu ( Prud’homme et al., 2020 ). Znižovala tiež inzulitídu, zatiaľ čo protilátka blokujúca Klotho mala opačný účinok. Tieto zistenia boli podobné ako u iných, ktorí používali metódy prenosu génov ( Lin a Sun, 2015a , b ). In vitro Klotho stimuloval prežitie, replikáciu a sekréciu inzulínu u ľudských β-buniek ( Prud’homme et al., 2017a ). Tiež inhiboval aktiváciu NF-KB, čo by mohlo vysvetliť antiapoptotický účinok. Klotho môže tiež chrániť β bunky prostredníctvom antioxidačných mechanizmov, ktoré môžu byť indukované napríklad Nrf2 a FoxO ( obrázok 3 ). Všimnite si, že hoci GABA bola aplikovaná iba pri predklinickom diabete, existuje množstvo GABAergických liekov v klinickom použití ( Palma et al., 2017 ) na liečbu epilepsie a iných chorôb. Na rozdiel od systémovo aplikovanej GABA, ktorá je blokovaná BBB, mnohé z týchto liekov prekračujú túto bariéru a to môže spôsobiť viaceré nepriaznivé účinky. Okrem GABA nie je známe, či GABAergné lieky zvyšujú hladiny Klotho.
Paradoxne, myši s deficitom Klotho ( KL kl/kl ) sú hypoglykemické, napriek tomu, že majú nízku hladinu inzulínu ( Razzaque 2012 ). To pravdepodobne vyplýva zo skutočnosti, že Klotho potláča citlivosť na inzulín, a preto bunky Klotho-negatívnych myší lepšie reagujú na inzulín. To vyvoláva otázku, či je Klotho vhodným prostriedkom na liečbu cukrovky. Predpokladáme, že liečba diabetu Klotho bude užitočná, keď sa obnovia normálne plazmatické hladiny. Je zaujímavé, že nadmerná expresia Klotho u transgénnych myší vyvolala miernu inzulínovú rezistenciu a predĺžila životnosť ( Kurosu et al., 2005 ). V experimentálnych T1D a T2D, ako je uvedené v tomto prehľade, bola terapia Klotho prospešná. Či ľudia zareagujú podobne, nie je známe. V našej práci sa Klotho proteín podával v nízkej dávke každých 48 hodín ( Prud’homme et al., 2020 ). Má polčas iba 7 hodín ( Hu et al., 2016 ), čo je relevantné pre účinnosť a potenciálne nepriaznivé účinky.
Jednou z kľúčových otázok týkajúcich sa antidiabetických liekov, ako sú GABA a GLP-1, je, či ich všeobecné antidiabetické účinky (Wang Q, et al., 2019; Drucker, 2018 ) zosilňujú Klotho nepriamo, alebo či existuje priamejší vplyv na expresiu génu Klotho. Toto nebolo dôkladne študované, ale predpokladáme, že je to spôsobené kombináciou mechanizmov. Pri cukrovke vyvolanej STZ je STZ toxický pre obličky ( Cheng MF et al., 2010 ), nielen pankreatické β bunky, a znižuje Klotho v obehu a obličkách ( Prud’homme et al., 2017a ). Toto bolo takmer úplne zvrátené perorálnym podávaním GABA. Štúdie iných ukázali, že liečba GABA chráni obličky pred tubulárnou fibrózou a atrofiou v modeli subtotálnej nefrektómie ( Sasaki et al., 2007 ) a ischemicko-reperfúznym poškodením ( Kobuchi et al., 2009 ). To je v súlade so skutočnosťou, že renálne tubulárne bunky exprimujú GABA receptory ( Amenta et al., 1988 ; Sarang et al., 2008 ; Takano et al., 2014 ). GABA teda môže chrániť renálne tubulárne bunky pred poškodením, takže pokračujú v produkcii Klotho na takmer normálnych hladinách. Avšak GABA a/alebo GLP-1RA priamo indukovali expresiu Klotho v pankreatických p bunkách. Je zaujímavé, že promótor Klotho má transkripčné väzbové miesto pre PAX4, čo je transkripčný faktor nevyhnutný na diferenciáciu β buniek v pankrease ( Sosa-Pineda et al., 1997 ; Xu a Sun, 2015 ). GLP-1 indukuje expresiu PAX4 v ľudských ostrovčekoch ( Brun et al., 2008 ), a preto to môže (spolu s ďalšími faktormi) indukovať expresiu Klotho.
Okrem cukrovky sa zistilo, že Klotho zohráva úlohu pri obezite. U ľudí sú hladiny Klotho v CSF znížené u obéznych subjektov ( Landry et al., 2021a , b ). Intracerebroventrikulárna injekcia Klotho u myší náchylných na diabetes znížila príjem potravy, zlepšila homeostázu glukózy a znížila telesnú hmotnosť ( Landry et al., 2020 ). Súviselo to so signalizáciou FGFR v neurónoch. Štúdie týchto autorov ukázali, že Klotho pôsobí na oblúkové jadro hypotalamu (ARC) s cieľom regulovať metabolizmus ( Landry et al., 2021b ). Neuróny aj astrocyty boli cielené a bola zdokumentovaná signalizácia FGFR prostredníctvom PI3K (neuróny) alebo ERK (astrocyty). Je zaujímavé, že nezistili žiadnu koreláciu medzi systémovými koncentráciami Klotho a CSF a mechanizmus regulácie Klotho v CSF zostáva nejasný. Zdá sa, že cirkulujúci Klotho tiež hrá dôležitú úlohu v metabolizme. Systémová liečba Klotho u myší znížila adipozitu a akumuláciu lipidov v pečeni; a zvýšený výdaj chudej hmoty a energie (Rao Z, et al., 2019). Pri liečbe Klotho odhalila kvantitatívna PCR nižšiu expresiu lipogénnych génov.
8 Klotho ako supresor nádorov
Existuje značný dôkaz, že Klotho je molekula supresoru nádorov, ako je uvedené inde ( Rubinek a Wolf, 2016 ; Abolghasemi a kol., 2019 ; Sachdeva a kol., 2020 ; Ewendt a kol., 2021 ). To je v súlade s jeho schopnosťou inhibovať dráhy spojené s rakovinou, vrátane Wnt, IGF-1, TGF-p a NF-KB, ako bolo uvedené vyššie ( obrázok 3 ; tabuľka 1 ). Expresia Klotho je utlmená alebo umlčaná u takmer všetkých vyšetrených typov rakoviny. Klotho-pozitívne nádory majú vo všeobecnosti lepšiu prognózu. Zdá sa, že k supresii Klotho dochádza rovnakým mechanizmom ako pri iných tumor supresorových génoch. Zahŕňa to napríklad hypermetyláciu DNA v miestach promótora, modifikácie histónov a miRNA ( Rubinek a Wolf, 2016 ; Abolghasemi et al., 2019 ). V experimentálnych modeloch nádorov má Klotho protinádorové účinky in vivo a/alebo in vitro . Toto bolo preukázané pri rakovine prsníka ( Wolf a kol., 2008 ; Ligumsky a kol., 2015 ), rakovine pankreasu ( Abramovitz a kol., 2011 ) a iných rakovinách vrátane rakoviny hrubého čreva, žalúdka a hepatocelulárneho karcinómu (HCC). Napríklad pri rakovine pankreasu bol Klotho slabo exprimovaný v nádore ( Abramovitz et al., 2011 ). Nadmerná expresia Klotho alebo terapia s-Klotho inhibovala rast rakovinových buniek pankreasu. Viaceré štúdie skutočne ukázali, že Klotho inhibuje proliferáciu rakovinových buniek, tvorbu kolónií a inváziu; a podporuje apoptózu a autofágiu ( Sachdeva et al., 2020 ). Podobne in vivo nútená nadmerná expresia Klotho v nádore alebo systémové podávanie s-Klotho inhibovalo rast niekoľkých typov nádorov. Dôležité je, že to zahŕňa ľudské nádory xenoštepované do myší. Membránovo viazané aj rozpustné formy Klotho majú supresorové účinky na nádory.
9 Klotho a neurodegeneratívne choroby
Nízke hladiny Klotho korelujú s neurodegeneratívnym ochorením a kognitívnym poškodením ( Vo a kol., 2018 ; Hanson a kol., 2021 ), ako aj s krehkosťou ( Veronesi a kol., 2021 ). Napríklad u obyvateľov domovov dôchodcov bola nízka hladina Klotho v sére spojená so zlým poznaním, krehkosťou, závislosťou a častými pádmi ( Sanz et al., 2021 ). Úloha Klotho v mozgu nie je dobre pochopená, ale existujú dôkazy, že má neuroprotektívny účinok. V centrálnom nervovom systéme je Klotho produkovaný choroidálnym plexom a nachádza sa v CSF. Je tiež široko exprimovaný v mozgu, prevažne v oblastiach šedej hmoty vrátane hipokampu ( Clinton a kol., 2013 ; Cararo-Lopes a kol., 2017 ). Produkujú ho neuróny aj oligodendrocyty. Expresia Klotho v mozgu je znížená u starnúcich subjektov a skorej Alzheimerovej choroby ( Fung et al., 2022 ). V myšacích modeloch Alzheimerovej choroby je nadmerná expresia Klotho prospešná.
Morfologicky je pri Alzheimerovej chorobe mozog charakterizovaný nadbytkom β-amyloidných (Ap) plakov, ako aj neurónovou agregáciou Tau proteínu tvoriacou neurofibrilárne klbká (NFT) ( Thal et al., 2013 ). Bolo navrhnuté, že tieto látky sú toxické pre neuróny. Klotho prepožičiava neurónovú rezistenciu voči oxidačnému a endoplazmatickému retikulovému stresu a predpokladá sa, že chráni pred toxicitou Ap a NFT ( Zeldich a kol., 2014 ; Fung a kol., 2022 ). Ap je generovaný proteolytickým štiepením amyloidového prekurzorového proteínu (APP) sekretázami ( Lichtenthaler et al., 2022 ). APP je transmembránový proteín typu 1 a jeho štiepenie α-sekretázou má za následok uvoľnenie (odlupovanie) veľkého extracelulárneho segmentu, ktorý netvorí amyloid. Tento proces štiepenia sa uskutočňuje primárne pomocou ADAM10 a je podobný generácii s-Klotho. Avšak spracovanie APP ß-sekretázou a y-sekretázou generuje malé sekretované Ap peptidy, ktoré tvoria amyloid. Je zaujímavé, že štiepenie a-sekretázy zabraňuje následnej tvorbe amyloidogénnych peptidov ( Lichtenthaler et al., 2022 ). Preto môže byť prospešné zvýšenie aktivity α-sekretázy alebo zníženie aktivity β- a y-sekretázy.
Jeden z možných mechanizmov ochorenia zahŕňa nedostatočnú autofágiu, čo je proces, ktorý normálne degraduje a odstraňuje proteíny a iné látky, čím bráni ich toxickej akumulácii v bunke. Klotho zlepšuje autofágiu a to môže chrániť pred neurodegeneratívnymi stavmi, ako je Alzheimerova choroba ( Fung et al., 2022 ). Zdá sa, že Klotho tiež prispieva k produkcii oligodendrocytického myelínu a udržiavaniu integrity bielej hmoty. Existujú dôkazy o zápalovej zložke Alzheimerovej choroby, ako aj niektorých ďalších neurodegeneratívnych ochorení, ktoré môžu zahŕňať aktiváciu NF-KB ( Ju wang et al., 2019 ). Tak, ako na iných miestach, Klotho môže mať prospešné protizápalové a antioxidačné účinky v mozgu. Klotho môže podporovať diferenciáciu mikroglií na protizápalový typ (M2) namiesto zápalového typu (M1) ( Fung et al., 2022 ).
Hipocampus hrá hlavnú úlohu v pamäti. Je to jedna z mála oblastí mozgu, ktorá si zachováva neurogenézu v dospelosti a pri Alzheimerovej chorobe dochádza k strate neurónov v tejto oblasti ( Rao YL, et al., 2022 ). Je pozoruhodné, že nedávna práca naznačuje, že Klotho stimuluje neurogenézu v hipokampe. Bolo to evidentné napríklad pri porovnaní myší s deficitom Klotho oproti myšiam s nadmernou expresiou Klotho ( Laszczyk et al., 2017 ). V súlade so stratou neurónov vykazujú myši s deficitom Klotho závažné kognitívne defekty. Nedávno sa ukázalo, že prerušovaný pôst u myší zvyšuje expresiu Klotho a neurogenézu v hipokampe ( Dias et al., 2021 ). Ukázalo sa, že prerušované hladovanie je lepšie ako obmedzenie kalórií a bolo spojené so zlepšením dlhodobej pamäte. Klotho-deficientné myši nereagovali na prerušované hladovanie. Je možné, že lieková terapia môže dosiahnuť podobné výsledky. Napríklad na potkanom modeli metabolického syndrómu inhibítor DPP-4 vildagliptín (zvyšuje GLP-1) zvýšil hipokampálny Klotho, súčasne znížil zápalové a apoptotické biomarkery v tomto mieste a zabránil neurodegeneratívnym zmenám ( Yossef et al., 2020 ). To zlepšilo pamäť u liečených potkanov. Všimnite si, že väčšina inhibítorov DPP-4, vrátane vildagliptínu, neprechádza hematoencefalickou bariérou (BBB), ale zvyšuje cirkulujúci GLP-1 ( Deacon, 2019 ), ktorý prechádza. Na potkanom modeli kognitívneho poškodenia vyvolaného STZ simvastatín zvýšil hipokampálny Klotho a zlepšil kognitívnu funkciu ( Adeli et al., 2017 ). Podobne iní zistili, že liečba rosiglitazónom zlepšila cerebrálnu expresiu Klotho ( Chen LJ, et al., 2014 ). V poslednej štúdii bola prospešná aj intracerebroventrikulárna infúzia rekombinantného Klotho. Alternatívne niektorí autori dodali Klotho génovým prenosom ( tabuľka 2 ), ako je napríklad intracerebroventrikulárna injekcia lentivírusového vektora kódujúceho Klotho ( Zeng et al., 2019 ). Vyššie uvedené zistenia týkajúce sa Klotho pri neurodegeneratívnych patológiách boli všetky získané u hlodavcov a zostáva určiť, či sú použiteľné pre ľudí.
Pokiaľ je nám známe, terapia na báze Klotho nebola aplikovaná u ľudí s neurodegeneratívnymi ochoreniami. Napriek tomu, v porovnaní s jedincami s nízkymi hladinami Klotho, tí s vysokými hladinami (CSF alebo plazma) majú lepšie kognitívne funkcie a nižší výskyt demencie ( Semba et al., 2014 ; Shardell et al., 2016 ). Predpokladalo sa, že hladiny Klotho v krvi a CSF sú nezávislé. Na rozdiel od tohto názoru niektorí výskumníci nedávno uviedli, že hladiny v sére a CSF silne korelujú a vysoké hladiny predpovedajú lepšiu kognitívnu funkciu ( Kundu et al., 2022 ). Toto je veľmi zaujímavé pre budúce klinické štúdie, najmä pokiaľ ide o lieky alebo iné liečby, ktoré zvyšujú Klotho. Výhradou je, že dôvody nezrovnalostí medzi štúdiami nie sú jasné, pokiaľ ide o hladiny Klotho v CSF alebo obeh. Existuje veľa obáv o špecifickosť niektorých anti-Klotho protilátok používaných v ELISA a iných testoch ( Kuro-o, 2019 ). Niektorí autori nedávno skúmali túto otázku vo vzorkách ľudského séra od pacientov so širokým rozsahom funkcie obličiek ( Neyra et al., 2020 ). Toto sa uskutočnilo široko používaným komerčným testom ELISA, ako aj imunoprecipitačným imunoblotovaním (IP-IB) s vysoko špecifickou protilátkou Klotho. Výsledky IP-IB Klotho korelovali s funkciou obličiek oveľa lepšie ako ELISA. Cykly zmrazovania a rozmrazovania zhoršili detekciu Klotho. Tieto zistenia naznačujú, že spoliehanie sa výlučne na test ELISA by mohlo byť zavádzajúce a manipulácia so vzorkou je dôležitá. Ďalším relevantným pozorovaním je široká variabilita hladín Klotho medzi jednotlivcami. Napríklad vo vekovej skupine 18 – 35 rokov mala približne polovica jednotlivcov nízke hladiny, porovnateľné so staršími subjektmi, zatiaľ čo druhá polovica mala vysoké hladiny ( Espuch-Oliver et al., 2022 ). Vo vekovej skupine 55 – 85 rokov však boli hladiny stále nižšie. Hladiny CSF sú tiež dosť variabilné ( Kundu et al., 2022 ). Vzhľadom na to by sa výsledky získané z malých skupín mali interpretovať opatrne. Biologický význam tejto variability je nejasný a sú potrebné dlhodobé štúdie.
Zdá sa, že variant génu Klotho označený KL-VS (2 aminokyselinové substitúcie v proteíne) poskytuje určitú ochranu pred Alzheimerovou chorobou, ale iba v heterozygotnom stave ( Driscoll a kol., 2021 ; Nietzel a kol., 2021 ). Priaznivé účinky boli pozorované len v niektorých podskupinách, ktoré boli študované. Spôsob účinku tohto variantu je nejasný, aj keď je zaujímavé, že sa zaznamenalo zníženie akumulácie Tau.
9.1 Súhrn referencií pre relevantnosť choroby
Pozri tiež tabuľku 1 . Obličky: Cheng a kol., 2010 ; Kuro-o, 2010 , 2019 , 2021 ; Maquigussa a kol., 2018 ; Oishi a kol., 2021 ; Prud’homme a kol., 2017a , 2020 ; Saar-Kovrov a kol., 2021 ; Takenaka a kol., 2017 ; Xing a kol., 2021 ; Zou a kol., 2018 . Kardiovaskulárne: Chen K, a kol., 2021 ; Freunlich a kol., 2021 ; Manrique a kol., 2016 ; Myung a kol., 2022 ; Martin-Nunez a kol., 2022 ; Olejnik a kol., 2018 , 2020 ; Tyrenkov a kol., 2021 ; Wang F a Zheng, 2022 ; Yang a kol., 2015 . Mozog: Dias a kol., 2021 ; Driscoll a kol., 2021 ; Cararo-Lopes a kol., 2017 ; Fung a kol., 2022 ; Hanson a kol., 2021 ; Kundu a kol., 2022 ; Laszczyk a kol., 2017 ; Mýtych, 2022 ; Nietzel a kol., 2021; Sanz a kol., 2021 . Rakovina: Doi a kol., 201 ; Ewendt a kol., 2021 ; Rubinek a vlk, 2016 ; Sachdeva a kol., 2020 . Pľúca: Huang Q. a kol., 2020; Gao a kol., 2015 . Kosti: Kuro-o, 2017 . 2019, 2021; Tasnim a kol., 2021 ; Metabolizmus a diabetes: Keles a kol., 2016 ; Landry a kol., 2021a , b ; Lin a Sun, 2012 , 2015a , 2015b ; Nie a kol., 2017 ; Prud’homme a kol., 2017a , 2020 ; Razzaque, 2012 ; Takenaka a kol., 2019 .
10 prístupov k terapii založenej na Klotho
10.1 Lieky v klinickom použití pre rôzne stavy
Vzhľadom na výhody Klotho proti starnutiu a viacerým chorobám, ako sa tu uvádza v predklinických modeloch ( obrázok 4 ; tabuľka 1 ), je veľmi zaujímavé nájsť uplatnenie u ľudí. U ľudí je dobre známe, že plazmatické hladiny Klotho klesajú s vekom a pri bežných chorobách, ako sú chronické ochorenia obličiek, cukrovka a neurodegeneratívne ochorenia. Vzhľadom na to by bolo samozrejmým a bezpečným cieľom obnovenie normálnych hladín. Niektoré lieky už boli spomenuté pre ich potenciálne použitie pri špecifických ochoreniach. V mnohých hlásených štúdiách ( Tabuľka 2 a 3 ) bolo hlavným účinkom zvýšenie hladín Klotho, ktoré sa približovalo alebo rovnalo normálu, a nie nad normálne. Na základe štúdií na myšiach môže nadmerná expresia predĺžiť životnosť, ale bezpečnosť je menej istá, pretože dôležité by mohli byť iné faktory (možno inzulínová rezistencia). Výhradou je, že prakticky všetky dostupné informácie o liečbe na báze Klotho boli získané od hlodavcov.
Prekvapivo veľký počet terapií zvyšuje hladiny Klotho v experimentálnych modeloch ( tabuľky 2 a 3 ), buď v obehu alebo špecifických orgánoch. Niektorým zahrnutým zlúčeninám sa v literatúre venovala značná pozornosť ako geroprotektívnym molekulám ( Soo et al., 2020 ), ale zvyčajne s nedostatočným preskúmaním ich kapacity indukcie Klotho. To zahŕňa niekoľko liekov, ktoré sa v súčasnosti klinicky používajú na rôzne stavy ( tabuľka 2 ).
10.1.1 Inhibítory RAAS
Inhibítory RAAS sú najlepšie zdokumentované klinické lieky zvyšujúce Klotho; najmä losartan a valsartan, ktoré blokujú receptor angiotenzínu II (AT1). V skutočnosti ide o niektoré z mála liekov, u ktorých sa v klinických štúdiách preukázalo, že zlepšujú plazmatický Klotho ( Karalliedde a kol., 2013 ; Lim SC a kol., 2014 ; Janic a kol., 2019 ). Napríklad losartan zvýšil hladiny Klotho o 23 % u diabetických pacientov ( Lim SC et al., 2014 ). To je v súlade so schopnosťou angiotenzínu II potlačiť expresiu Klotho ( Yoon HE, 2011 ). Nie je známe, či augmentácia Klotho prispieva ku klinickej účinnosti týchto liekov. Je potrebné poznamenať, že tieto výsledky boli získané u diabetických subjektov a zostáva určiť, či lieky budú pôsobiť podobne pri iných ochoreniach alebo u zdravých subjektov.
10.1.2 Statíny
Statíny sú ďalším typom často aplikovaných liekov, ktoré zvyšujú Klotho ( tabuľka 2 ); z klinických štúdií však existujú obmedzené dôkazy. Najzaujímavejšie je, že podávanie nízkodávkovej kombinácie fluvastatínu a valsartanu zvýšilo expresiu Klotho a SIRT1 , ktoré sú obe spojené so zvýšenou dlhovekosťou ( Janic et al., 2019 ). Naopak, mTOR a NF-KB neboli zvýšené. Statíny a inhibítory RAAS sú veľmi bežnou kombináciou liekov v medicíne a tento typ liečby môže mať účinok proti starnutiu. Pretože tieto lieky majú veľký vplyv na krvný tlak, lipidy, funkciu obličiek, kardiovaskulárne ochorenia a ďalšie faktory, bude ťažké určiť špecifický prínos Klotho.
10.1.3 Pentoxifylín
V klinickej štúdii sa ukázalo, že pentoxifylín zvyšuje Klotho ( Navarro-Gonzalez et al., 2018 ). Tento liek sa zvyčajne používa na liečbu ischemickej choroby periférnych ciev, existujú však aj iné indikácie. U diabetických pacientov (typ 2) sa zvýšil sérový a močový Klotho, ale zmena sérových hladín bola relatívne mierna (~ 6 %). Pacienti mali pokročilé ochorenie obličiek, čo môže vysvetliť nízku odpoveď. V klinických štúdiách je hlavným faktorom, ktorý treba zvážiť, zdravie obličiek. Renálna atrofia (typicky nefroskleróza) môže byť závažná u pacientov s cukrovkou alebo hypertenziou, a pretože väčšina cirkulujúceho Klotho pochádza z obličiek, môže to znížiť hladiny. Nízke hladiny Klotho sú teda pravdepodobne odrazom ochorenia obličiek a ak je objem obličiek veľmi nízky, potom akákoľvek liečba liekom nemusí Klotho zvýšiť.
10.1.4 Vitamín D
Suplementácia vitamínu D tiež zvyšuje Klotho, ako sa ukázalo u detí s chronickým ochorením obličiek ( Lerch et al., 2018 ). Deti s miernym až stredne ťažkým ochorením obličiek mali na začiatku štúdie znížené hladiny Klotho v sére, čo sa normalizovalo liečbou vitamínom D. Na rozdiel od toho pacienti so závažným ochorením obličiek mali nezmenené hladiny Klotho. To predstavuje obmedzené údaje dostupné z klinickej práce. U hlodavcov však existuje jasný dôkaz o posilnení Klotho vitamínom D ( Tabuľka 2 ). Na vysvetlenie tohto terapeutického účinku je potrebné zvážiť komplexné interakcie, ktoré sú základom homeostázy FGF23/Klotho/vitamínu D, ako bolo nedávno preskúmané ( Haussler a kol., 2020 ; Neyra a kol., 2021 ).
10.1.5 Rapamycín a Everolimus
Rapamycín (sirolimus) a everolimus sú inhibítory mTOR v klinickom použití, u ktorých sa ukázalo, že oba zvyšujú Klotho ( tabuľka 2 ). Rapamycín bol dlho navrhovaný ako geroprotektívne liečivo. Klinicky sa rapamycín aplikuje predovšetkým ako imunosupresívny liek na prevenciu odmietnutia transplantovaného orgánu. Liečba príjemcov obličkového transplantátu rapamycínom vyvolala hypofosfatémiu a inzulínovú rezistenciu ( Tataranni et al., 2011 ). Ošetrenie proximálnych renálnych tubulárnych buniek rapamycínom in vitro vyvolalo expresiu Klotho. Autori navrhli, že Klotho sprostredkúval klinické zmeny súvisiace s rapamycínom. V klinickej štúdii po transplantácii obličky everolimus významne zvýšil hladiny Klotho v sére ( Mizusaki et al., 2019 ). Títo výskumníci merali hladiny Klotho pred transplantáciou a 1 rok po nej. Porovnávali tiež pacientov, ktorí boli liečení everolimom, s pacientmi, ktorí neboli liečení. V oboch prípadoch boli hladiny Klotho posilnené týmto liekom. Je zaujímavé, že samotná transplantácia obličky zvýšila Klotho, v súlade s nefrektómiou znižujúcou Klotho, a ďalším dôkazom, že oblička je hlavným zdrojom cirkulujúceho Klotho ( Hu et al., 2016 ). Zvýšené hladiny Klotho u pacientov liečených everolimom významne nekorelovali so zlepšením funkcie obličiek, vrátane sérového kreatinínu, dusíka močoviny v krvi, eGFR, vápnika a fosforu. Táto štúdia bola malá a súbežne sa podávali iné imunosupresívne látky, čo obmedzuje interpretáciu. U potkanov s chronickým zlyhaním obličiek perorálna liečba rapamycínom tiež zvýšila klotho, čo zlepšilo vaskulárnu kalcifikáciu ( Zhao et al., 2015 ). Výhody sa pripisovali Klotho a inhibícii mTOR na základe množstva farmakologických a genetických metód. Je pozoruhodné, že rapamycín nemohol zabrániť vaskulárnemu ochoreniu, keď bola expresia Klotho potlačená siRNA alebo u myší s knockoutom Klotho. Tieto štúdie poukazujú na rapamycín a everolimus ako účinné lieky na zvýšenie hladín Klotho. Tieto lieky však môžu mať značné nepriaznivé účinky, vrátane zníženej imunity voči infekčným agens a bunkovej toxicity v niektorých orgánoch. Napríklad rapamycín je toxický pre pankreatické β bunky ( Prud’homme et al., 2013 ).
10.1.6 Antidiabetiká
Ukázalo sa, že niekoľko klinických antidiabetík zvyšuje expresiu Klotho u myší, napr. GLP-1RA, inhibítory DPP-4, metformín a agonisty receptorov y (PPAR-y) aktivovaných peroxizómovým proliferátorom ( Tabuľka 2 ). Niektoré boli spomenuté už skôr. Metformín, ktorý je pravdepodobne najčastejšie predpisovaným antidiabetikom pre T2D, zvýšil Klotho v obehu, obličkách a moči ( Xue et al., 2019 ). Znižoval hladiny mTOR a tento účinok bol zvrátený potlačením Klotho. Metformín bol dlho navrhovaný ako geroprotektívne liečivo, hoci existujú protichodné zistenia ( Mohammed et al., 2021 ). V prípade liečby cukrovky je varovaním, že metformín zvyšuje citlivosť na inzulín, zatiaľ čo Klotho má opačný účinok.
PPAR-y je transkripčný faktor regulujúci citlivosť na inzulín a adipogenézu a agonistické lieky nachádzajú uplatnenie pri cukrovke a poruchách lipidov. Agonisty PPAR-y sú ďalšou triedou liečiv, ktoré zvyšujú expresiu Klotho ( tabuľka 2 ). V 5′-lemujúcej oblasti ľudského génu Klotho existuje nekanonický prvok reagujúci na PPAR (Zhang H, et al., 2008), ktorý pravdepodobne vysvetľuje túto aktivitu. Títo autori ukázali, že in vitro a/alebo in vivo troglitazón, ciglitazón a rosiglitazón zvýšili expresiu Klotho mRNA. Pozorovali zvýšený proteín Klotho v obličkách. Antagonista PPAR-y mal opačné účinky.
10.1.6.1 Súhrn referencií pre klinické lieky, ktoré zvyšujú Klotho
Pozri tiež tabuľku 2 . Inhibítory renín-angiotenzín-aldosterón: Janic et al., 2019 ; Li X a Zhou, 2010 ; Lim SC, a kol., 2014 ; Yoon a kol., 2011 . Statíny: Adeli et al., 2017 ; Janic a kol., 2019 ; Kuwahara a kol., 2008 ; Narumiya a kol., 2004 ; Yoon a kol., 2012 . Agonisty PPAR-y: Chen LJ, et al., 2014 ; Cheng a kol., 2017 ; Huang KC, a kol., 2014 ; Maquigussa a kol., 2018 ; Shen a kol., 2018 ; Zhang H, a kol., 2008. Inhibítory mTOR: Mizusaki a kol., 2019 ; Tabibzadeh, 2021 ; Tataranni a kol., 2011 ; Zhao Y, a kol., 2015. Vitamín D: Haussler a kol., 2020 ; Kuro-o, 2019 ; Lerch a kol., 2018 ; Tsujikawa al., 2003. Antidiabetiká: Manrique et al., 2016 ; Liu W, a kol., 2017, 2021; Son a kol., 2019 ; Wang Q, a kol., 2019 ; Yossef a kol., 2020 ; Xue a kol., 2019 . Pentoxifylín: Navarro-Gonzalez a kol., 2018 . Iné: Zhou a kol., 2022 ; Kang a Xu, 2016 .
10.2 Lieky v predklinickom vývoji
Množstvo liečiv, rekombinantných proteínov (alebo peptidov) a génových terapií bolo testovaných na predklinických modeloch chorôb na ich schopnosť zvyšovať hladiny Klotho ( tabuľka 3 ). Špecifické príklady podávania rekombinantného s-Klotho alebo peptidov boli diskutované skôr. Génová terapia sa uskutočňovala s vírusovými vektormi alebo plazmidmi a bunková terapia sa tiež javí ako uskutočniteľná ( Franco et al., 2021 ). V niektorých prípadoch, ako sú rekombinantné proteínové a génové terapie, sú pravdepodobne limitujúce faktory realizovateľnosť, potenciálne nepriaznivé účinky, dostupnosť a vysoké náklady.
Rekombinantná proteínová terapia Klotho bola úspešne aplikovaná na liečbu obličkových, kardiovaskulárnych a neurodegeneratívnych ochorení, ako aj cukrovky a rakoviny ( tabuľka 3 ). Vo všeobecnosti išlo o krátkodobú štúdiu s injekciou s-Klotho, domény KL1 alebo relevantných peptidov. Nízke dávky sú zvyčajne účinné. Rekombinantný proteín bol dodávaný systémovo alebo do komôr mozgu pri neurologických ochoreniach.
Bežné zložky potravy/diety (napr. astaxantín, kurkumín a resveratrol) a tradičné lieky a/alebo ich extrahované zlúčeniny (napr. baikalín, cordycepín, ženšen a ligustilid) boli preskúmané a úspešne indukovali Klotho ( tabuľka 3 ). Niektoré z nich sú často súčasťou voľne predajných nutraceutík. Vo všeobecnosti tieto zlúčeniny zlepšili alebo obnovili hladiny Klotho v modeloch chorôb hlodavcov, kde sú depresívne. Táto práca bola vykonaná na zvieracích modeloch a nie je známe, či tieto látky môžu zvýšiť Klotho u ľudí.
10.3 Cvičenie a kondícia
Najzaujímavejšie je, že fyzické cvičenie a šport sú jednoduchými spôsobmi na zvýšenie cirkulujúceho Klotho, ako hodnotili iní ( Amaro-Gahete et al., 2018 ). Jediný záchvat cvičenia prechodne zvýši cirkulujúci Klotho ( Tan a kol., 2018 ; Morishima a Ochi, 2021 ). Štúdia FIT-AGEING skúmala súvislosť medzi fyzickou aktivitou a kondíciou a plazmatickými hladinami s-Klotho u dospelých sedavých ľudí v strednom veku ( Amaro-Gahete et al., 2019a , b ). Autori uviedli, že vyššia aktivita a kondícia sú spojené so zvýšenými hladinami s-Klotho. Hodnotili účinky rôznych kurzov cvičenia počas 12-týždňového obdobia a zistili, že všetky typy zvýšili Klotho v porovnaní so žiadnym cvičením. Uvádzajú, že vyššie s-Klotho sa spája so znížením tukovej hmoty a zvýšením chudej hmoty.
10.3.1 Súhrn referencií pre lieky alebo liečby vo vývoji
Pozri tiež tabuľku 3 . agonista GABA receptora: Liu W, et al., 2021 ; Prud’homme a kol., 2017a ; Son a kol., 2019 ; Wang Q, a kol., 2019 . Rekombinantný proteín Klotho alebo peptidy: Ali et al., 2020 ; Chen TH, a kol., 2013 ; Doi a kol., 2011 ; Gupta a kol., 2022 ; Hu a kol., 2017 ; Leon a kol., 2017 ; Maltese a kol., 2017 ; Mencke a kol., 2017 ; Myung a kol., 2022 ; Oh a kol., 2018 ; Prud’homme a kol., 2020 ; Rao Z, a kol., 2019 ; Takenaka a kol., 2018 , 2019 , 2020 ; Yang a kol., 2015 ; Yuan a kol., 2022 ; Zhong a kol., 2020 . Génová terapia: Arbel Rubinstein a kol., 2021 ; Franco a kol., 2021 ; Lin & Sun, 2015a , b ; Ni a kol., 2021 ; Mencke a kol., 2017 ; Shin a kol., 2019 ; Xiang a kol., 2021 ; Zeng a kol., 2019 . Tradičné lieky, nutraceutiká a doplnky: Dehghani et al., 2019 ; Feng a Huang, 2022 ; Hsu a kol., 2014 ; Kamel a kol., 2022 ; Long a kol., 2018 ; Li SS, a kol., 2018 ; Lim SW, a kol., 2019 ; Long a kol., 2018 ; Ostojič a Engeset, 2021 ; Zhang P, a kol., 2016 ; Zhang XT, a kol., 2020 ; Zhou a kol., 2015 . Cvičenie a šport: Amaro-Gahete et al., 2018 , 2019a , b ; Morishima a Ochi, 2021 ; Tan a kol., 2018 .
10.4 BBB a liečba neurodegeneratívnych chorôb
BBB (krvno-mozgová bariéra) blokuje vstup väčšiny liekov do mozgu a predstavuje výzvu pre terapiu neurodegeneratívnych stavov, ako sa uvádza v prehľade ( Partridge, 2012 ; Nilles et al., 2022 ). Niektoré malé liečivá rozpustné v tukoch prechádzajú difúziou sprostredkovanou lipidmi.
Iné lieky môžu byť prepravované špecifickými transportérmi. Veľké liečivá (zvyčajne proteíny) sa vo všeobecnosti nekrížia alebo sa neprenášajú veľmi neefektívne, ale prenos možno zlepšiť pomocou inžinierskych konštruktov pre transport sprostredkovaný receptormi. Okrem toho sa zdá, že neuropilín-1 uľahčuje vstup do mozgu niektorých bunkových penetračných peptidov, väčších molekúl alebo vírusov ( Balistreri a kol., 2021 ; Prud’homme a Glinka, 2012 ; Ruoslahti, 2017 ; Zhao L, a kol. ., 2021 ).
Zaujímavým príkladom sú lieky blokujúce RAAS, pretože ich možno rozdeliť na typy BBB-crossing a non-crossing. Napríklad blokátory AT1-receptora losartan, irbesartan, olemesartan a eprosartan neprenikajú cez BBB, zatiaľ čo valsartan, telmisartan a kandesartan sa krížia ( Ho et al., 2017 ). Pamäť sa zlepšila u starších dospelých liečených blokátormi AT1 a užívatelia liekov krížiacich sa s BBB dosahovali lepšie výsledky ako tí, ktorí užívali lieky bez kríženia ( Ho et al., 2017 ). Subjekty, ktoré dostávali lieky s krížením BBB, mali pri zobrazovaní tiež výrazne menej hyperintenzít bielej hmoty (WMH). Iní výskumníci ( Ohrui et al., 2004 ) hlásili podobné zistenia u pacientov, ktorí dostávali BBB-kríženie inhibítorov angiotenzín-konvertujúceho enzýmu (ACE). Tí, ktorí boli liečení liekmi krížiacimi sa s BBB (kaptopril, perindopril), mali výrazne nižšie riziko vzniku Alzheimerovej choroby v porovnaní s tými, ktorí užívali lieky, ktoré sa neskrížili (imidapril, enalapril). Ochranný mechanizmus je nejasný. Môžu lieky inhibujúce RAAS krížiace sa s BBB zvýšiť Klotho v mozgu alebo CSF? Pokiaľ je nám známe, o ľuďoch neexistujú žiadne údaje. V cerebri spontánne hypertenzných potkanov však liečba valsartanom znížila poškodenie buniek, čo bolo určené ultraštrukturálnymi zmenami, ako je tvorba apoptotického tela ( Li a Zhou, 2010 ). Liečba valsartanom bola spojená so zvýšenou intracerebrálnou expresiou Klotho, ako sa stanovilo pomocou RT-PCR, imunohistochémie a Western blottingu. To poskytuje určitý dôkaz, aj keď obmedzený, že valsartan môže zvýšiť expresiu Klotho v mozgu. Predpokladá sa, že statíny majú aspoň nejaké neuroprotektívne účinky. Môžu byť tiež rozdelené na BBB-kríženie a nekríženie. Lipofilné statíny (napr. atorvastatín, lovastatín, simvastatín) sa krížia a hydrofilné nie ( Wood et al., 2010 ). V tomto prípade simvastatín zvýšil Klotho v hipokampe potkanov ( Adeli et al., 2017 ). Je teda možné, ale ešte nie isté, že tak inhibítory RAAS, ako aj statíny môžu zvýšiť expresiu Klotho v ľudskom mozgu, čo by mohlo chrániť pred neurodegeneratívnymi ochoreniami.
Niektoré antidiabetiká (alebo aspoň členovia týchto rodín liekov), ktoré zvyšujú hladiny cirkulujúceho Klotho u hlodavcov, môžu prechádzať cez BBB. Patria sem niektoré GLP-1RA ( Pujadas a Drucker, 2016 ), metformín ( Moreira, 2014 ) a aspoň jeden inhibítor DPP-4 (omariglipín) ( Ayoub et al., 2018 ). Inhibítory DPP-4 zvyšujú GLP-1 ( Deacon, 2019 ), prirodzený GLP-1RA, ktorý prechádza cez BBB. Tieto antidiabetiká možno u niektorých pacientov zlepšujú neurodegeneratívne ochorenie ( Mehan et al., 2022 ), ale potenciálny prínos Klotho, pokiaľ je nám známe, nebol preskúmaný.
Ako je uvedené v tabuľke 3 , ukázalo sa, že niekoľko zlúčenín odvodených z tradičných liekov alebo iných zdrojov zvyšuje expresiu Klotho u hlodavcov. Je zaujímavé, že niektoré z týchto zlúčenín prechádzajú cez BBB, ako napríklad astaxantín, baikalín, cordycepín, ligustilid a resveratrol. Zdá sa, že kurkumín ( Reddy et al., 2018 ), ako aj zložka ženšenu, ginsenosid RG1 ( Zhao YN, et al., 2018 ), aspoň do určitej miery prechádzajú cez BBB. Ligustilid je mimoriadne zaujímavý, pretože ľahko prechádzal cez BBB a chránil u myší pred ischemickým poranením mozgu a Alzheimerovou chorobou. V ischemickom mozgu sa znížil NF-κB a zápalové zmeny ( Long et al., 2018 ). V modeli Alzheimerovej choroby bola inhibovaná signalizácia IGF-1/Akt/mTOR ( Kuang et al., 2014 ). Ligustilid zvýšil Klotho v choroidálnom plexe. Znížená regulácia Klotho znížila ochranné účinky. Zdá sa, že neuroprotekcia závisí aspoň čiastočne od zvýšenia aktivity ADAM10. Ako bolo uvedené vyššie, štiepenie a-sekretázou APP pomocou ADAM10 produkuje uvoľnenie veľkého nepatogénneho segmentu a zabraňuje tvorbe malých peptidov tvoriacich Ap ( Lichtenthaler et al., 2022 ). Liečba ligustilidom zvýšila aktivitu ADAM10, čo malo za následok vyššiu produkciu s-Klotho aj neamiloidogénneho APP. Tvorba Ap plakov bola znížená u myší liečených ligustilidom. Autori ( Kuang et al., 2014 ) dospeli k záveru, že zlepšenie s-Klotho a α-spracovanie APP prispeli k zlepšeniu neurodegeneratívneho ochorenia.
V niektorých prípadoch môžu lieky, ktoré neprechádzajú cez BBB, nepriamymi mechanizmami zlepšiť Alzheimerovu chorobu alebo iné neurodegeneratívne stavy. Zlepšenie hypertenzie, hyperlipidémie, cukrovky alebo funkcie obličiek môžu mať veľký vplyv na mozog. U mladých a starých myší systémové podávanie peptidu odvodeného od Klotho, ktorý neprechádza cez BBB, zlepšilo kogníciu a znížilo neurodegeneratívnu patológiu ( Leon et al., 2017 ). Iní ukázali, že systémové dodávanie buď s-Klotho alebo iba domény KL1 tiež zlepšilo kogníciu ( Gupta et al., 2022 ). Nie je jasné, ako sú tieto prospešné účinky na báze Klotho sprostredkované, ale penetrácia do mozgu sa nezdá byť potrebná, a to si zaslúži ďalšie skúmanie.
11 Záver a budúce smerovanie
V tomto rukopise sme preskúmali mnohé aspekty biológie Klotho. Najlepšie charakterizované aspekty sa týkajú fyziológie obličiek a najmä homeostázy fosfátov a vápnika. Dôležitosť interakcie FGF23/Klotho/FGFR bola evidentná u myší s deficitom Klotho. Hlavnými výsledkami boli hyperfosfatémia a hypervitaminóza D. Syndróm zrýchleného starnutia u týchto myší sa výrazne zlepšil nízkofosfátovou diétou. Zriedkavé mutácie Klotho alebo FGF23 u ľudí vedú k hyperfosfatemickému syndrómu, ktorý sa vyznačuje masívnou kalcifikáciou tkaniva a systémovým zápalom. Pri chronickom ochorení obličiek rôznej etiológie, ako aj pri cukrovke, je produkcia Klotho znížená. Predpokladalo sa, že klesajúca funkcia obličiek a súvisiaca nízka hladina Klotho urýchľujú alebo zhoršujú mnohé z patológií vyskytujúcich sa v starobe. To môže zahŕňať napríklad precipitáciu CPP v tkanive ( Kuro-o, 2021 ).
Zdá sa nepravdepodobné, že všetky prejavy Klotho insuficiencie súvisia so signalizáciou indukovanou FGF23. V skutočnosti, ako je uvedené v niekoľkých častiach, Klotho blokuje hlavné cesty, ktoré zohrávajú úlohu pri starnutí. To sa týka špecificky inhibície TGF-p, Wnt, IGF-1 a NF-KB. Sú to zložité dráhy, ktoré ovplyvňujú obrovské množstvo biologických procesov. Bez ohľadu na iné mechanizmy, ktoré môžu viesť k starnutiu, je teda pravdepodobne dôležitá nadmerná aktivácia týchto dráh. Toto je najzreteľnejšie pri fibróze, zápale a rakovine, ktorým Klotho bráni. Okrem toho Klotho aktivuje antioxidačné dráhy, ako sú Nrf2 a FOXO, o ktorých sa predpokladá, že sprostredkovávajú účinky proti starnutiu.
Pokiaľ ide o budúce smerovanie, existuje niekoľko dôležitých ciest. Klotho je opísaná ako molekula proti starnutiu, aj keď neexistujú žiadne priame dôkazy, že spomaľuje starnutie u ľudí. Toto je jedna z najdôležitejších otázok, ktorú treba riešiť, ale bude si to vyžadovať veľmi dlhodobé štúdie. Klotho môže byť marker choroby (obličky, mozog alebo iné) alebo terapeutická molekula. Najmä funkcia (funkcie) Klotho v mozgu nie je dobre definovaná, ale sú zrejmé neuroprotektívne účinky. Hladiny Klotho v obehu alebo CSF by mohli naznačovať náchylnosť na ochorenie. Zdá sa, že viaceré lieky zvyšujú Klotho, ale štúdie na ľuďoch sú nedostatočné. Prispieva Klotho k terapeutickým účinkom niektorých liekov používaných na liečbu hypertenzie, hyperlipidémie, cukrovky alebo odmietnutia transplantátu? Mohli by niektoré z týchto liekov zlepšiť neurodegeneratívne ochorenia zlepšením Klotho? Toto všetko je neznáme. V skutočnosti sa okrem predklinickej práce prakticky neuskutočnili žiadne klinické výskumy liečby súvisiacej s liekom Klotho. Ďalším dôležitým aspektom je úloha Klotho ako nádorovej supresorovej molekuly. Toto je dobre zdokumentované na zvieracích modeloch, ale je potrebné ho vyvinúť v klinickom kontexte. Úzke klinické aplikácie výskumu Klotho možno vysvetliť obmedzeným pochopením jeho mechanizmov účinku, najmä mimo obličiek. Poznatky o molekulárnych a fyziologických aspektoch Klotho však v posledných rokoch značne pokročili, ako je uvedené v tomto prehľade.
Pre terapiu je realizovateľných niekoľko prístupov, ako je opísané v tomto rukopise. Klinický výskum zameraný na ktorýkoľvek z týchto prístupov by bol dôležitým príspevkom v tejto oblasti. Najpriamejším prístupom je podávanie rekombinantného Klotho alebo peptidov. Vzhľadom na relatívne krátky polčas rozpadu týchto produktov existuje potreba vyrábať dlhodobo pôsobiace konštrukty alebo formulácie s pomalým uvoľňovaním. Existuje len málo dôkazov z predklinických prác, že podávanie Klotho je toxické, aspoň pri obnovení normálnych hladín. Dlhodobé účinky nadnormálnych hladín však nie sú známe. Génová terapia je tiež uskutočniteľná, ale oveľa náročnejšia. Okrem niektorých liekov na klinický predpis existujú aj bežne dostupné voľnopredajné lieky, doplnky stravy alebo nutraceutiká, ktoré zvyšujú Klotho. Toto je založené na práci zvierat a bolo by veľmi zaujímavé preskúmať možnosť vylepšenia Klotho u ľudí. V prípade neurodegeneratívnych ochorení je dôležité, že niektoré lieky, vrátane niektorých tradičných liekov, prechádzajú cez BBB a môžu zvýšiť Klotho v mozgu. Napokon, cvičenie a dlhodobé udržiavanie kondície sú jednoduché metódy na zvýšenie Klotho. Na záver, väčšina štúdií o Klotho bola vykonaná na zvieracích modeloch chorôb a na zavedenie terapie Klotho na kliniku je potrebné vykonať značné množstvo práce. Tento prehľad poskytuje rámec mechanizmov a zistení, ktoré uprednostňujú klinické skúmanie liečby na báze Klotho.
Autorské príspevky
GP napísal rukopis, MK vykonal experimenty súvisiace s Klotho a pripravil obrázky pre tento rukopis, QW dohliadal na niektoré štúdie a upravoval rukopis. Všetci autori skontrolovali a schválili obsah tohto rukopisu.
Financovanie
Práca vykonaná autormi v tomto rukopise bola financovaná Medzinárodnou nadáciou pre výskum diabetu mladých ľudí (čísla grantu 2-SRA-2015-64-QR a 2-SRA-2018-497-AB), Nadáciou Nemocnice sv. Michala (Toronto , Kanada), Keenan Research Center for Biomedical Science (Toronto, Kanada), Canadian Diabetes Association (číslo grantu OG-3-13-4066) a Čínska národná vedecká nadácia (čísla grantov 81570518, 81630020, 81800751).
Konflikt záujmov
Autor QW bol zamestnaný v spoločnosti Shanghai Yinuo Pharmaceutical Co., Ltd. QW je držiteľom patentov súvisiacich s GLP-1.
Zostávajúci autori vyhlasujú, že výskum bol vykonaný bez akýchkoľvek obchodných alebo finančných vzťahov, ktoré by mohli byť chápané ako potenciálny konflikt záujmov.
Poznámka vydavateľa
Všetky tvrdenia vyjadrené v tomto článku sú výlučne tvrdeniami autorov a nemusia nevyhnutne predstavovať tvrdenia ich pridružených organizácií alebo tvrdenia vydavateľa, redaktorov a recenzentov. Žiadny produkt, ktorý môže byť hodnotený v tomto článku, alebo tvrdenie, ktoré môže uviesť jeho výrobca, nie je zaručené ani schválené vydavateľom.
Poďakovanie
Autori ďakujú Dr. Yelena Glinka, Dr. Anna Toulina, Dr. Wenjuan Liu a Dr. Dong Ok Son za ich odbornú prácu a rady pri vykonávaní experimentov z našich laboratórií. Niektoré obrázky na obrázkoch boli z obrázku Servier Medical Art ( smart.servier.com ), získaného na základe licencie Creative Commons Attribution 3.0 Unported License.
Referencie
Aashaq, S., Batool, A., Mir, SA, Beigh, MA, Andrabi, KI a Shah, ZA (2022). Signalizácia TGF‐β: Rekapitulácia ciest nezávislých od SMAD a závislých od SMAD. J. Cell. Fyziológia 237 (1), 59–85. doi:10.1002/jcp.30529
Abolghasemi, M., Yousefi, T., Maniati, M. a Qujeq, D. (2019). Súhra Klotho so signálnou dráhou a mikroRNA pri rakovine. J Cell. Biochem. 120 (9), 14306–14317. doi:10.1002/jcb.29022
Abramovitz, L., Rubinek, T., Ligumsky, H., Bose, S., Barshack, I., Avivi, C., et al. (2011). Vnútorné opakovanie KL1 sprostredkúva aktivity supresora nádoru Klotho a inhibuje signalizáciu bFGF a IGF-I pri rakovine pankreasu. Clin. Cancer Res. 17 (13), 4254-4266. doi:10.1158/1078-0432.CCR-10-2749
Adeli, S., Zahmatkesh, M., Tavoosidana, G., Karimian, M. a Hassanzadeh, G. (2017). Simvastatín zlepšuje hipokampálne klotho u potkanieho modelu kognitívneho poklesu vyvolaného streptozotocínom. Prog. Neuro-Psychopharmacology Biol. Psychiatria 72, 87–94. doi:10.1016/j.pnpbp.2016.09.009
Ali, MF, Venkatarayappa, SKB, Benny, M., Rojas, C., Yousefi, K., Shehadeh, LA, et al. (2020). Účinky suplementácie Klotho na poškodenie obličiek vyvolané hyperoxiou v modeli hlodavcov postnatálnej nefrogenézy. Pediatr. Res. 88, 565-570. doi:10.1038/s41390-020-0803-z
Amaro-Gahete, FJ, De-la-O, A., Jurado-Fasoli, L., Espuch-Oliver, A., de Haro, T., Gutierrez, A., et al. (2019b2019b). Cvičenie zvyšuje plazmatické hladiny S-Klotho u sedavých dospelých v strednom veku: Randomizovaná kontrolovaná štúdia. Štúdia FIT-AGEING. J. Sports Sci. 37 (19), 2175-2183. doi:10.1080/02640414.2019.1626048
Amaro-Gahete, FJ, de-la-O, A., Jurado-Fasoli, L., Gutiérrez, Á., Ruiz, JR, Castillo, MJ, et al. (2019a). Asociácia fyzickej aktivity a kondície s plazmatickými hladinami S-Klotho u dospelých v sedavom veku v strednom veku: Štúdia FIT-AGEING. Maturita 123, 25-31. doi:10.1016/j.maturitas.2019.02.001
Amaro-Gahete, FJ, de-la-O, A., Jurado-Fasoli, L., Ruiz, JR, Castillo, MJ a Gutiérrez, Á. (2019a2018). Úloha cvičenia pri regulácii proteínu S-Klotho: Systematický prehľad. Cas 11 (2), 100-107. doi:10.2174/1874609811666180702101338
Amenta, F., Cavallotti, C., Iacopino, L. a Erdó, S. a L. (1988). Autorádiografická lokalizácia agonistu [3H]-muscimolu GABAA receptora v obličkách potkana. Farmakológia 36 (6), 390–395. doi:10.1159/000138327
Andersen, B., Omar, BA, Rakipovski, G., Raun, K. a Ahrén, B. (2015). Fibroblastový rastový faktor 21 zabraňuje zhoršeniu glykémie v modeloch diabetu u myší s deficitom inzulínu. Eur. J. Pharmacol. 764, 189–194. doi:10.1016/j.ejphar.2015.07.003
Andrukhova, O., Zeitz, U., Goetz, R., Mohammadi, M., Lanske, B. a Erben, RG (2012). FGF23 pôsobí priamo na proximálne tubuly obličiek na vyvolanie fosfatúrie prostredníctvom aktivácie signálnej dráhy ERK1/2-SGK1. Kosť 51 (3), 621–628. doi:10.1016/j.bone.2012.05.015
Anor, R., Andrukhova, O., Ritter, E., Zeitz, U. a Erben, RG (2012). Klotho nemá fyziologickú úlohu nezávislú od vitamínu D v homeostáze glukózy, kostnom obrate a sekrécii PTH v ustálenom stave in vivo . PLoS One 7 (2), e31376. doi:10.1371/journal.pone.0031376
Arbel Rubinstein, T., Reuveni, I., Hesin, A., Klein-Goldberg, A., Olauson, H., Larsson, TE, a kol. (2021). Transgénny model odhaľuje úlohu Klotho pri rozvoji rakoviny pankreasu a pripravuje cestu pre novú terapiu založenú na Klotho. Cancers 13 (24), 6297. doi:10.3390/cancers13246297
Ayoub, BM, Mowaka, S., Safar, MM, Ashoush, N., Arafa, MG, Michel, HE a kol. (2018). Zmena polohy omarigliptínu ako intranazálneho antiparkinsoník raz týždenne. Sci. Rep. 8 (1), 8959. doi:10.1038/s41598-018-27395-0
Baba, AB, Rah, B., Bhat, GR, Mushtaq, I., Parveen, S., Hassan, R., a kol. (2022). Signalizácia transformujúceho rastového faktora beta (TGF-β) pri rakovine – zrada vo vnútri. Predné. Pharmacol. 13, 791272. doi:10.3389/fphar.2022.791272
Balistreri, G., Yamauchi, Y. a Teesalu, T. (2021). Rozšírený mechanizmus vstupu vírusu: Interakcia motívu C-konca a neuropilínového receptora. Proc. Natl. Akad. Sci. USA 118 (49), e2112457118. doi:10.1073/pnas.2112457118
Ben-Dov, IZ, Galitzer, H., Lavi-Moshayoff, V., Goetz, R., Kuro-o, M., Mohammadi, M., et al. (2007). Prištítne telieska sú cieľovým orgánom pre FGF23 u potkanov. J. Clin. Vyšetrovať. 117 (12), 4003-4008. doi:10.1172/JCI32409
BonDurant, LD, a Potthoff, MJ (2018). Fibroblastový rastový faktor 21: Všestranný regulátor metabolickej homeostázy. Annu. Rev. Nutr. 38, 173-196. doi:10.1146/annurev-nutr-071816-064800
Brun, T., He, KHH, Lupi, R., Boehm, B., Wojtusciszyn, A., Sauter, N., et al. (2008). Transkripčný faktor Pax4 spojený s diabetom je exprimovaný v ľudských pankreatických ostrovčekoch a je aktivovaný mitogénmi a GLP-1. Hum. Mol. Genet. 17 (4), 478-489. doi: 10,1093/hmg/ddm325
Buchanan, S., Combet, E., Stenvinkel, P. a Shiels, PG (2020). Klotho, starnutie a zlyhávajúca oblička. Predné. Endocrinol. (Lausanne) 11, 560. doi:10.3389/fendo.2020.00560
Buendía, P., Carracedo, J., Soriano, S., Madueño, JA, Ortiz, A., Martín-Malo, A., et al. (2015). Klotho zabraňuje translokácii NFκB a chráni endotelové bunky pred starnutím vyvolaným urémiou. Gerona 70 (10), 1198–1209. doi:10.1093/gerona/glu170
Buendía, P., Ramírez, R., Aljama, P. a Carracedo, J. (2016). Klotho zabraňuje translokácii NFκB. Vitam. Horm. 101, 119-150. doi:10.1016/bs.vh.2016.02.005
Bzowska, M., Jura, N., Lassak, A., Black, RA a Bereta, J. (2004). Nádorový nekrotický faktor-a stimuluje expresiu TNF-a konvertujúceho enzýmu v endotelových bunkách. Eur. J. Biochem. 271 (13), 2808–2820. doi:10.1111/j.1432-1033.2004.04215.x
Cararo-Lopes, MM, Mazucanti, CHY, Scavone, C., Kawamoto, EM a Berwick, DC (2017). Význam α-KLOTHO pre centrálny nervový systém: Niektoré kľúčové otázky. Ageing Res. 36 , 137–148. doi:10.1016/j.arr.2017.03.003
Chen, C.-D., Podvin, S., Gillespie, E., Leeman, SE a Abraham, CR (2007). Inzulín stimuluje štiepenie a uvoľňovanie extracelulárnej domény Klotho pomocou ADAM10 a ADAM17. Proc. Natl. Akad. Sci. USA 104 (50), 19796–19801. doi:10.1073/pnas.0709805104
Chen, G., Liu, Y., Goetz, R., Fu, L., Jayaraman, S., Hu, M.-C., a kol. (2018). α-Klotho je neenzymatický molekulárny skelet pre signalizáciu hormónu FGF23. Príroda 553 (7689), 461–466. doi:10.1038/nature25451
Chen, K., Wang, S., Sun, QW, Zhang, B., Ullah, M. a Sun, Z. (2021). Nedostatok Klotho spôsobuje starnutie srdca prostredníctvom narušenia dráhy Nrf2-GR. Circ. Res. 128 (4), 492-507. doi:10.1161/CIRCRESAHA.120.317348
Chen, L.-J., Cheng, M.-F., Ku, P.-M., a Lin, J.-W. (2014). Rosiglitazón zvyšuje cerebralklothoexpresiu na zvrátenie baroreflexu u diabetických potkanov typu 1. BioMed Res. Int. 2014, 1.–9. doi:10.1155/2014/309151
Chen, T.-H., Kuro-o, M., Chen, C.-H., Sue, Y.-M., Chen, Y.-C., Wu, H.-H., a kol. (2013). Vylučovaný proteín Klotho obnovuje retenciu fosfátov a potláča zrýchlené starnutie u mutantných myší Klotho. Eur. J. Pharmacol. 698 (1-3), 67-73. doi:10.1016/j.ejphar.2012.09.032
Cheng, L., Zhang, L., Yang, J. a Hao, L. (2017). Aktivácia receptora y aktivovaného peroxizómovým proliferátorom inhibuje vaskulárnu kalcifikáciu upreguláciou Klotho. Exp. Ther. Med. 13 (2), 467-474. doi:10.3892/etm.2016.3996
Cheng, M.-F., Chen, L.-J., and Cheng, J.-T. (2010). Zníženie Klotho v obličkách diabetických potkanov indukovaných streptozotocínom. J. Biomed. Biotechnol. 2010, 1.–7. doi:10.1155/2010/513853
Clinton, SM, Glover, ME, Maltare, A., Laszczyk, AM, Mehi, SJ, Simmons, RK, a kol. (2013). Expresia klotho mRNA a proteínu v mozgovom parenchýme potkanov od skorého postnatálneho vývoja do dospelosti. Brain Res. 1527, 1–14. doi:10.1016/j.brainres.2013.06.044
Coppede, F. (2021). Mutácie podieľajúce sa na syndrómoch predčasného starnutia. Tacg Vol. 14, 279-295. doi:10.2147/TACG.S273525
Dalton, GD, Xie, J., An, S.-W., and Huang, C.-L. (2017). Nové pohľady na mechanizmus pôsobenia rozpustného Klotho. Predné. Endocrinol. 8, 323. doi:10.3389/fendo.2017.00323
de Almeida, AJPO, de Almeida Rezende, MS, Dantas, SH, de Lima Silva, S., de Oliveira, JCPL, de Lourdes Assunção Araújo de Azevedo, F. a kol. (2020). Odhalenie úlohy zápalu a oxidačného stresu pri kardiovaskulárnych ochoreniach súvisiacich s vekom. Oxidative Med. Cell. Longev. 2020, 1.–20. doi:10.1155/2020/1954398
de Streel, G. a Lucas, S. (2021). Zacielenie imunosupresie pomocou TGF-p1 na imunoterapiu rakoviny. Biochem. Pharmacol. 192, 114697. doi:10.1016/j.bcp.2021.114697
Diakon, CF (2019). Fyziológia a farmakológia DPP-4 v glukózovej homeostáze a liečbe diabetu 2. typu. Predné. Endocrinol. 10, 80. doi:10.3389/fendo.2019.00080
Dehghani, A., Hafizibarjin, Z., Najjari, R., Kaseb, F. a Safari, F. (2019). Súbežné podávanie resveratrolu a 1,25-dihydroxyvitamínu D chráni srdce pred starnutím vyvolaným D-galaktózou u potkanov: hodnotenie sérových a srdcových hladín klotho. Aging Clin. Exp. Res. 31 (9), 1195-1205. doi:10.1007/s40520-018-1075-x
Derynck, R. a Budi, EH (2019). Špecifickosť, všestrannosť a kontrola signalizácie rodiny TGF-β. Sci. Signál. 12 (570), eaav5183. doi:10.1126/scisignal.aav5183
Dias, GP, Murphy, T., Stangl, D., Ahmet, S., Morisse, B., Nix, A., et al. (2021). Prerušovaný pôst zvyšuje dlhodobú konsolidáciu pamäte, hipokampálnu neurogenézu dospelých a expresiu génu dlhovekosti Klotho. Mol. Psychiatria 26 (11), 6365–6379. doi:10.1038/s41380-021-01102-4
Dinkova-Kostova, AT, Kostov, RV a Kazantsev, AG (2018). Úloha signalizácie Nrf2 v boji proti neurodegeneratívnym ochoreniam. FEBS J. 285 (19), 3576-3590. doi:10.1111/febs.14379
Doi, S., Zou, Y., Togao, O., Pastor, JV, John, GB, Wang, L. a kol. (2011). Klotho inhibuje signalizáciu transformujúceho rastového faktora-β1 (TGF-β1) a potláča renálnu fibrózu a metastázy rakoviny u myší. J. Biol. Chem. 286 (10), 8655-8665. doi:10.1074/jbc.M110.174037
Dolegowska, K., Marchelek-Mysliwiec, M., Nowosiad-Magda, M., Slawinski, M. a Dolegowska, B. (2019). Členovia podrodiny FGF19: FGF19 a FGF21. J. Physiol. Biochem. 75 (2), 229-240. doi:10.1007/s13105-019-00675-7
Donate-Correa, J., Martín-Núñez, E., Delgado, NP, de Fuentes, MM, Arduan, AO, Mora-Fernández, C., a kol. (2016). Dôsledky fibroblastového rastového faktora / a-Klotho systému v metabolizme glukózy a cukrovke. Cytokine Growth Factor Rev. 28, 71-77. doi:10.1016/j.cytogfr.2015.12.003
Driscoll, I., Ma, Y., Gallagher, CL, Johnson, SC, Asthana, S., Hermann, BP a kol. (2021). Záťaž Tau súvisiaca s vekom a kognitívne deficity sú u heterozygotov KLOTHO KL-VS oslabené. Jad 82 (3), 1369–1370. PMID: 34250957]. doi:10.3233/JAD-219006
Drucker, DJ (20182018). Mechanizmy účinku a terapeutická aplikácia glukagónu podobného peptidu-1. Cell. Metab. 27 (4), 740-756. doi:10.1016/j.cmet.2018.03.001
Erben, RG a Andrukhova, O. (2017). FGF23-Klotho signálna os v obličkách. Kosť 100, 62–68. doi:10.1016/j.bone.2016.09.010
Erben, RG (2018). Fyziologické účinky fibroblastového rastového faktora-23. Predné. Endocrinol. 9, 267. doi:10.3389/fendo.2018.00267
Espuch-Oliver, A., Vázquez-Lorente, H., Jurado-Fasoli, L., de Haro-Muñoz, T., Díaz-Alberola, I., López-Velez, M. d. S., a kol. (2022). Referencie Hodnoty sérových hladín rozpustného α-Klotho s použitím imunosorbentného testu viazaného na enzým u zdravých dospelých vo veku 18-85 rokov. Jcm 11 (9), 2415. doi:10,3390/jcm11092415
Ewendt, F., Feger, M. a Föller, M. (2021). Úloha fibroblastového rastového faktora 23 (FGF23) a aKlotho pri rakovine. Predné. Cell. Dev. Biol. 8, 601006. doi:10.3389/fcell.2020.601006
Feng, Y. a Huang, Q. (2022)., 22. PubMed , 433–440. doi:10.1111/ggi.14376Ochranné účinky cordycepínu proti starnutiu vyvolanému d‐galaktózou u potkanov: Pohľad zo srdca Geriatr. Gerontol. Int.
Fountoulakis, N., Maltese, G., Gnudi, L. a Karalliedde, J. (2018). Znížené hladiny anti-agingového hormónu Klotho predpovedajú pokles funkcie obličiek pri cukrovke 2. typu. J. Clin. Endocrinol. Metab. 103 (5), 2026–2032. doi:10.1210/jc.2018-00004
Franceschi, C., Garagnani, P., Morsiani, C., Conte, M., Santoro, A., Grignolio, A., et al. (2018). Kontinuum starnutia a chorôb súvisiacich s vekom: bežné mechanizmy, ale rôzne miery. Predné. Med. 5, 61. doi:10.3389/fmed.2018.00061
Franco, ML, Beyerstedt, S. a Rangel, É. B. (2021). Klotho a mezenchymálne kmeňové bunky: Prehľad bunkovej a génovej terapie pre chronické ochorenie obličiek a akútne ochorenie obličiek. Pharmaceutics 14 (1), 11. doi:10.3390/pharmaceutics14010011
Freundlich, M., Gamba, G. a Rodriguez-Iturbe, B. (2021). Fibroblastový rastový faktor 23-Klotho a hypertenzia: experimentálne a klinické mechanizmy. Pediatr. Nephrol. 36 (10), 3007-3022. doi:10.1007/s00467-020-04843-6
Frieling, J. a Roeder, T. (2020). Faktory, ktoré ovplyvňujú premenu diétnych obmedzení na dlhší život. IUBMB Life 72 (5), 814–824. doi:10.1002/iub.2224
Fulop, T., Larbi, A., Pawelec, G., Khalil, A., Cohen, AA, Hirokawa, K., et al. (2021). Imunológia starnutia: zrod zápalu. Clin. Allerg. Immunol. , 1-14. doi:10.1007/s12016-021-08899-6
Fung, TY, Iyaswamy, A., Sreenivasmurthy, SG, Krishnamoorthi, S., Guan, X.-J., Zhu, Z., a kol. (2022). Klotho stimulátor autofágie ako potenciálny terapeutický cieľ pre Alzheimerovu chorobu: prehľad. Biomedicines 10 (3), 705. doi:10.3390/biomedicines10030705
Gao, W., Guo, L., Yang, Y., Wang, Y., Xia, S., Gong, H. a kol. (2022). Rozoberanie presluchu medzi cestami odozvy Nrf2 a NF-KB pri toxicite vyvolanej drogami. Predné. Cell. Dev. Biol. 9, 809952. doi:10.3389/fcell.2021.809952
Gao, W., Yuan, C., Zhang, J., Li, L., Yu, L., Wiegman, CH, a kol. (2015). Expresia Klotho je znížená v epiteliálnych bunkách dýchacích ciest pri CHOCHP: účinky na zápal a oxidačné poškodenie. Clin. Sci. (Londýn) 129 (12), 1011–1023. doi:10.1042/CS20150273
Groppe, J., Hinck, CS, Samavarchi-Tehrani, P., Zubieta, C., Schuermann, JP, Taylor, AB, a kol. (2008). Kooperatívne zostavenie signálnych komplexov nadrodiny TGF-P je sprostredkované dvoma odlišnými mechanizmami a odlišnými spôsobmi väzby na receptor. Mol. Cell. 29 (2), 157-168. doi:10.1016/j.molcel.2007.11.039
Guo, Y., Zhuang, X., Huang, Z., Zou, J., Yang, D., Hu, X., et al. (2018). Klotho chráni srdce pred poškodením vyvolaným hyperglykémiou inaktiváciou zápalu sprostredkovaného ROS a NF-KB In vitro aj in vivo . Biochimica Biophysica Acta (BBA) – Mol. Basis Dis. 1864 (1), 238–251. doi:10.1016/j.bbadis.2017.09.029
Gupta, S., Moreno, AJ, Wang, D., Leon, J., Chen, C., Hahn, O., a kol. (2022). Pubmed , JN–RM. JN-RM-2458-21. doi:10.1523/JNEUROSCI.2458-21.2022KL1 doména faktora dlhovekosti klotho napodobňuje metabolóm kognitívnej stimulácie a zlepšuje kogníciu u mladých a starnúcich myší J. Neurosci.
Guthrie, G., Vonderohe, C. a Burrin, D. (2022). Expresia, regulácia a funkcia fibroblastového rastového faktora 15/19: Prehľad. Mol. Cell. Endocrinol. 548, 111617. doi:10.1016/j.mce.2022.111617
Haga, M. a Okada, M. (2022). Systémové prístupy na skúmanie úlohy NF-κB signalizácie pri starnutí. Biochem. J. 479 (2), 161-183. doi:10.1042/BCJ20210547
Hanson, K., Fisher, K. a Hooper, NM (2021). Využitie neuroprotektívnych účinkov α-klotho na riešenie kognitívnej dysfunkcie súvisiacej so starnutím a neurodegeneráciou. Neurónový signál 5 (2), NS20200101. doi:10.1042/NS20200101
Hart, AW, Baeza, N., Apelqvist, Å. a Edlund, H. (20002000). Zoslabenie signalizácie FGF v myších β-bunkách vedie k cukrovke. Nature 408 (6814), 864–868. doi:10.1038/35048589
Haussler, MR, Livingston, S., Sabir, ZL, Haussler, CA a Jurutka, PW (2020). Receptor vitamínu D sprostredkúva nespočetné množstvo biologických účinkov závislých od jeho 1,25‐ dihydroxyvitamínu D ligandu: Odlišné regulačné témy odhalené indukciou klotho a fibroblastového rastového faktora-23. JBMR Plus 5 (1), e10432. doi:10.1002/jbm4.10432
Hayat, R., Manzoor, M. a Hussain, A. (2022)., 46. PubMed , 863–877. doi:10.1002/cbin.11797Wnt signálna dráha: Komplexný prehľad Cell. Biol. Int.
Ho, JK, Nation, DA a Nation, Iniciatíva DAAlzheimer’s Disease Neuroimaging Initiative (2017). Pamäť je zachovaná u starších dospelých, ktorí užívajú blokátory AT1 receptorov. Alz Res. Ther. 9 (1), 33. doi:10.1186/s13195-017-0255-9
Hsu, S.-C., Huang, S.-M., Chen, A., Sun, C.-Y., Lin, S.-H., Chen, J.-S., a kol. (2014). Resveratrol zvyšuje expresiu génu Klotho proti starnutiu prostredníctvom signálnej dráhy sprostredkovanej aktivačným transkripčným faktorom 3/c-Jun komplexom. Int. J. Biochem. Cell. Biol. 53, 361-371. doi:10.1016/j.biocel.2014.06.002
Hu, MC, Bian, A., Neyra, J. a Zhan, M. (2015). Klotho, kmeňové bunky a starnutie. Cia 10, 1233–1243. [. doi:10.2147/CIA.S84978
Hu, MC, Kuro-o, M. a Moe, OW (2013). Klotho a chronické ochorenie obličiek. Contrib. Nephrol. 180, 47-63. doi:10.1159/000346778
Hu, MC, Shi, M., Gillings, N., Flores, B., Takahashi, M., Kuro-o, M. a kol. (2017). Rekombinantné a-Klotho môže byť profylaktické a terapeutické pre akútnu až chronickú progresiu ochorenia obličiek a uremickú kardiomyopatiu. Kidney Int. 91 (5), 1104-1114. doi:10.1016/j.kint.2016.10.034
Hu, MC, Shi, M., Zhang, J., Addo, T., Cho, HJ, Barker, SL, a kol. (2016). Renálna produkcia, príjem a manipulácia s cirkulujúcim αKlotho. J Am Soc Nephrol 27 (1), 79–90. doi:10.1681/ASN.2014101030
Huang, K.-C., Cherng, Y.-G., Chen, L.-J., Hsu, C.-T. a Cheng, J.-T. (2014). Rosiglitazón je účinný na zlepšenie poškodenia obličiek u diabetických potkanov typu 1. Horm. Metab. Res. 46 (4), 240-244. doi:10.1055/s-0033-1357161
Huang, Q., Chen, Y., Shen, S., Wang, Y., Liu, L., Wu, S., a kol. (2020). Klotho antagonizuje pľúcnu fibrózu potláčaním aktivácie, migrácie a produkcie extracelulárnej matrice pľúcnych fibroblastov: terapeutický význam pre idiopatickú pľúcnu fibrózu. Starnutie 12 (7), 5812–5831. doi:10.18632/starnutie.102978
Huang, T., David, L., Mendoza, V., Yang, Y., Villarreal, M., De, K., et al. (2011). Signalizácia TGF-β je sprostredkovaná dvoma autonómne fungujúcimi pármi TβRI:TβRII. EMBO J. 30 (7), 1263–1276. doi:10.1038/emboj.2011.54
Ito, N. a Fukumoto, S. (2021). Vrodené hyperfosfatemické stavy spôsobené nedostatočnou aktivitou FGF23. Calcif. Tissue Int. 108 (1), 104-115. doi:10.1007/s00223-020-00659-6
Janić, M., Lunder, M., Novaković, S., Škerl, P. a Šabovič, M. (2019). Expresia génov dlhovekosti indukovaná nízkou dávkou kombinácie fluvastatínu a valsartanu s potenciálom predchádzať/liečiť „poruchy súvisiace so starnutím“. Ijms 20 (8), 1844. doi:10.3390/ijms20081844
Javidnia, S., Cranwell, S., Mueller, SH, Selman, C., Tullet, JMA, Kuchenbaecker, K., et al. (2022). Mendelovské randomizačné analýzy implikujú biogenézu translačného aparátu v ľudskom starnutí. Genome Res. 32 (2), 258-265. doi:10.1101/gr.275636.121
Johnson, SC (2018). Snímanie živín, signalizácia a starnutie: Úloha IGF-1 a mTOR pri starnutí a chorobách súvisiacich s vekom. Podbunka. Biochem. 90, 49-97. doi:10.1007/978-981-13-2835-0_3
Ju Hwang, C., Choi, D.-Y., Park, MH, a Hong, JT (2019). NF-KB ako kľúčový mediátor zápalu mozgu pri Alzheimerovej chorobe. Cnsnddt 18 (1), 3–10. doi:10.2174/1871527316666170807130011
Kamel, SS, Baky, NAA, Karkeet, RM, Osman, AMM, Sayed-Ahmed, MM a Fouad, MA (2022). Astaxantín zoslabuje inhibíciu expresie aldehyddehydrogenázy a proteínu Klotho v modeli akútneho kardiomyopatického potkana vyvolaného cyklofosfamidom. Clin. Exp. Pharma Physio 49 (2), 291-301. doi:10.1111/1440-1681.13598
Kang, W.-L. a Xu, G.-S. (2016). Atrasentan zvýšil expresiu klotho sprostredkovaním miR-199b-5p a zabránil poškodeniu renálnych tubulov pri diabetickej nefropatii. Sci. Rep. 6, 19979. doi:10.1038/srep19979
Kapahi, P., Kaeberlein, M. a Hansen, M. (2017). Diétne obmedzenia a dĺžka života: Lekcie z modelov bezstavovcov. Ageing Res. Zj 39, 3–14. [PMID: 28007498]. doi:10.1016/j.arr.2016.12.005
Karalliedde, J., Maltese, G., Hill, B., Viberti, G. a Gnudi, L. (2013). Účinok blokády renín-angiotenzínového systému na rozpustný Klotho u pacientov s diabetom 2. typu, systolickou hypertenziou a albuminúriou. Clin J Am Soc Nephrol 8 (11), 1899–1905. doi:10.2215/CJN.02700313
Keles, N., Dogan, B., Kalcik, M., Caliskan, M., Keles, NN, Aksu, F., et al. (2016). Je sérum Klotho ochranné proti ateroskleróze u pacientov s diabetes mellitus 1. typu? J. Diabetes jeho komplikácia. 30 (1), 126-132. doi:10.1016/j.jdiacomp.2015.09.013
Kilkenny, DM, a Rocheleau, JV (2016). Signalizačný komplex receptora FGF21. Vitam. Horm. 101, 17-58. doi:10.1016/bs.vh.2016.02.008
Kinoshita, S. a Kawai, M. (2016). Regulačná sieť FGF23/KLOTHO a jej úlohy pri ľudských poruchách. Vitam. Horm. 101, 151-174. doi:10.1016/bs.vh.2016.02.001
Kobuchi, S., Shintani, T., Sugiura, T., Tanaka, R., Suzuki, R., Tsutsui, H., et al. (2009). Renoprotektívne účinky kyseliny y-aminomaslovej na poškodenie obličiek vyvolané ischémiou/reperfúziou u potkanov. Eur. J. Pharmacol. 623 (1-3), 113-118. doi:10.1016/j.ejphar.2009.09.023
Kresovich, JK a Bulka, CM (2021). Nízke sérové klotho spojené s mortalitou zo všetkých príčin medzi národne reprezentatívnou vzorkou dospelých Američanov. J. Gerontol. A Biol. Sci. Med. Sci. 77, glab308452–456. doi:10.1093/gerona/glab308
Kuang, X., Chen, Y.-S., Wang, L.-F., Li, Y.-J., Liu, K., Zhang, M.-X., a kol. (2014). Upregulácia Klotho prispieva k neuroprotekcii ligustilidu v myšom modeli Alzheimerovej choroby. Neurobiol. Vek 35 (1), 169–178. doi:10.1016/j.neurobiolaging.2013.07.019
Kundu, P., Zimmerman, B., Quinn, JF, Kaye, J., Mattek, N., Westaway, SK, a kol. (2022)., 86. PubMed , 1471–1481. doi:10.3233/JAD-215719Hladiny α-Klotho v sére korelujú s hladinami mozgovomiechového moku a predpovedajú miery kognitívnej funkcie Jad
Kuro-O, M. (2018). Molekulárne mechanizmy, ktoré sú základom zrýchleného starnutia v dôsledku defektov v systéme FGF23-Klotho. Int. J. Nephrol. 2018, 9679841. doi:10.1155/2018/9679841
Kuro-o, M. (2010). Potenciálne spojenie medzi fosfátmi a starnutím – Poučenie od myší s deficitom Klotho. Mech. Starnutie Dev. 131 (4), 270-275. doi:10.1016/j.mad.2010.02.008
Kuro-o, M. (2021). Klotho a kalciproteínové častice ako terapeutické ciele proti zrýchlenému starnutiu. Clin. Sci. (Londýn) 135 (15), 1915–1927. doi:10.1042/CS20201453
Kuro-o, M. (2012). Klotho v zdraví a chorobe. Curr. Opin. Nephrol. Hypertenzia. 21 (4), 362-368. doi:10.1097/MNH.0b013e32835422ad
Kuro-o, M., Matsumura, Y., Aizawa, H., Kawaguchi, H., Suga, T., Utsugi, T. a kol. (1997). Mutácia myšacieho génu klotho vedie k syndrómu pripomínajúcemu starnutie. Príroda 390 (6655), 45–51. doi:10.1038/36285
Kuro-o, M. (2017). Systém FGF23 a Klotho mimo minerálneho metabolizmu. Clin. Exp. Nephrol. 21 (Suppl. 1), 64-69. doi:10.1007/s10157-016-1357-6
Kuro-o, M. (2019). Proteíny Klotho v zdraví a chorobe. Nat. Nephrol. 15 (1), 27-44. doi:10.1038/s41581-018-0078-3
Kurosu, H., Yamamoto, M., Clark, JD, Pastor, JV, Nandi, A., Gurnani, P., a kol. (2005). Potlačenie starnutia u myší hormónom Klotho. Veda 309 (5742), 1829–1833. doi:10.1126/science.1112766
Kuwahara, N., Sasaki, S., Kobara, M., Nakata, T., Tatsumi, T., Irie, H., et al. (2008). Inhibícia HMG-CoA reduktázy zlepšuje expresiu klotho proteínu proti starnutiu a artériosklerózu u potkanov s chronickou inhibíciou syntézy oxidu dusnatého. Int. J. Cardiol. 123 (2), 84–90. PMID: 17434618]. doi:10.1016/j.ijcard.2007.02.029
Landry, T., Laing, BT, Li, P., Bunner, W., Rao, Z., Prete, A., et al. (2020). Centrálne α-Klotho potláča aktivitu neurónov NPY/AgRP a reguluje metabolizmus u myší. Diabetes 69 (7), 1368–1381. doi:10.2337/db19-0941
Landry, T., Li, P., Shookster, D., Jiang, Z., Li, H., Laing, BT, a kol. (2021b). Centrálne cirkulujúce a-klotho nepriamo koreluje s ľudskou obezitou a moduluje oblúkovité bunkové populácie u myší. Mol. Metab. 44, 101136. doi:10.1016/j.molmet.2020.101136
Landry, T., Shookster, D. a Huang, H. (2021a). Cirkulujúce α-klotho reguluje metabolizmus prostredníctvom odlišných centrálnych a periférnych mechanizmov. Metabolizmus 121, 154819. doi:10.1016/j.metabol.2021.154819
Laszczyk, AM, Fox-Quick, S., Vo, HT, Nettles, D., Pugh, PC, Overstreet-Wadiche, L. a kol. (2017). Klotho reguluje postnatálnu neurogenézu a chráni pred stratou priestorovej pamäte súvisiacou s vekom. Neurobiol. Vek 59, 41–54. doi:10.1016/j.neurobiolaging.2017.07.008
Leon, J., Moreno, AJ, Garay, BI, Chalkley, RJ, Burlingame, AL, Wang, D., a kol. (2017). Periférna elevácia fragmentu Klotho zvyšuje funkciu a odolnosť mozgu u mladých, starnúcich a transgénnych myší s a-synukleínom. Cell. Rep. 20 (6), 1360–1371. doi:10.1016/j.celrep.2017.07.024
Lerch, C., Shroff, R., Wan, M., Rees, L., Aitkenhead, H., Kaplan Bulut, I., et al. (2018). Účinky výživovej suplementácie vitamínu D na markery metabolizmu kostí a minerálov u detí s chronickým ochorením obličiek. Nephrol. Vytáčať. Transpl. 33 (12), 2208-2217. doi:10.1093/ndt/gfy012
Li, L., Wang, Y., Gao, W., Yuan, C., Zhang, S., Zhou, H., a kol. (2015). Klotho redukcia v alveolárnych makrofágoch prispieva k zápalu vyvolanému extraktom z cigaretového dymu pri chronickej obštrukčnej chorobe pľúc. J. Biol. Chem. 290 (46), 27890–27900. doi:10.1074/jbc.M115.655431
Li, S.-s., He, A.-1., Deng, Z.-y., a Liu, Q.-f. (2018). Ginsenoside-Rg1 chráni pred fibrózou obličiek reguláciou signálnej dráhy Klotho/TGF-β1/Smad u potkanov s obštrukčnou nefropatiou. Biol. Pharm. Bull. 41 (4), 585-591. doi:10.1248/bpb.b17-00934
Li, X. a Zhou, Q. (2010). Účinok valsartanu na ultraštruktúru v mozgovom tkanive a expresiu génu Klotho u spontánne hypertenzných potkanov. Zhong Nan Da Xue Xue Bao Yi Xue Ban. 35 (3), 254-261. doi:10.3969/j.issn.1672-7347.2010.03.011
Li, X., Lu, P., Shao, X.-F., Jiang, T., Liu, F. a Li, G. (2021). Klotho reguluje prechod z epitelu na mezenchým vo Vitrovii Wnt/β-katenínovú dráhu a zmierňuje chronickú dysfunkciu aloštepu v modeli transplantácie obličky u potkanov. Ann. Transpl. 26, e930066. doi:10.12659/AOT.930066
Lichtenthaler, SF, Tschirner, SK a Steiner, H. (2022). Sekretázy pri Alzheimerovej chorobe: Nové poznatky o proteolýze APP a TREM2. Curr. Opin. Neurobiol. 72, 101-110. doi:10.1016/j.conb.2021.09.003
Ligumsky, H., Rubinek, T., Merenbakh-Lamin, K., Yeheskel, A., Sertchook, R., Shahmoon, S., et al. (2015). Nádorová supresorová aktivita Klotho pri rakovine prsníka je odhalená analýzou štruktúry a funkcie. Mol. Cancer Res. 13 (10), 1398-1407. doi:10.1158/1541-7786.MCR-15-0141
Lim, K., Groen, A., Molostvov, G., Lu, T., Lilley, KS, Snead, D., a kol. (2015). a-Klotho expresia v ľudských tkanivách. J. Clin. Endocrinol. Metabolizmus 100 (10), E1308–E1318. doi:10.1210/jc.2015-1800
Lim, SC, Liu, J.-J., Subramaniam, T. a Sum, CF (2014). Zvýšené cirkulujúce alfa-klotho blokátorom receptora angiotenzínu II losartanom je spojené so znížením albuminúrie u pacientov s diabetom 2. typu. J. Renin Angiotenzín Aldosterón Syst. 15 (4), 487-490. doi:10.1177/1470320313475905
Lim, SW, Shin, YJ, Luo, K., Quan, Y., Cui, S., Ko, EJ, a kol. (2019). Ženšen zvyšuje expresiu Klotho pomocou FoxO3-sprostredkovanej superoxiddismutázy mangánu v myšom modeli poškodenia obličiek vyvolaného takrolimom. Starnutie 11 (15), 5548–5569. doi:10.18632/starnutie.102137
Lin, W., Wu, X., Wen, J., Fei, Y., Wu, J., Li, X. a kol. (2021). Nikotínamid si zachováva expresiu Klotho a zmierňuje akútne poškodenie obličiek vyvolané rabdomyolýzou. Výživa 91-92, 111376. doi:10.1016/j.nut.2021.111376
Lin, Y. a Sun, Z. (2015b2015). Antiaging Gene Klotho zmierňuje apoptózu pankreatických β-buniek pri diabete 1. typu. Diabetes 64 (12), 4298-4311. doi:10.2337/db15-0066
Lin, Y. a Sun, Z. (2012). Antiaging Gene Klotho zvyšuje glukózou indukovanú sekréciu inzulínu zvýšením hladiny TRPV2 v plazmatickej membráne v MIN6 β-bunkách. Endokrinológia 153 (7), 3029–3039. doi:10.1210/en.2012-1091
Lin, Y. a Sun, Z. (2015a2015). In vivo pankreatická β-bunková špecifická expresia antiagingového génu Klotho: Nový prístup na ochranu β-buniek pri diabete 2. typu. Diabetes 64 (4), 1444–1458. doi:10.2337/db14-0632
Lindberg, K., Amin, R., Moe, OW, Hu, M.-C., Erben, RG, Östman Wernerson, A., et al. (2014). Obličky sú hlavným orgánom sprostredkujúcim klotho efekty. Clin J Am Soc Nephrol 25 (10), 2169–2175. doi:10.1681/ASN.2013111209
Liu, H., Fergusson, MM, Castilho, RM, Liu, J., Cao, L., Chen, J., et al. (2007). Rozšírená signalizácia Wnt v cicavčom modeli zrýchleného starnutia. Science 317 (5839), 803–806. doi:10.1126/science.1143578
Liu, W., Lau, HK, Son, DO, Jin, T., Yang, Y., Zhang, Z., et al. (2021). Kombinované použitie GABA a sitagliptínu podporuje proliferáciu ľudských β-buniek a znižuje apoptózu. J. Endocrinol. 248 (2), 133–143. doi:10.1530/JOE-20-0315
Liu, W., Son, DO, Lau, HK, Zhou, Y., Prud’homme, GJ, Jin, T. a kol. (2017). Kombinované perorálne podávanie GABA a inhibítora DPP-4 zabraňuje poškodeniu beta buniek a podporuje regeneráciu beta buniek u myší. Predné. Pharmacol. 8, 362. doi:10.3389/fphar.2017.00362
Long, F.-Y., Shi, M.-Q., Zhou, H.-J., Liu, D.-L., Sang, N. a Du, J.-R. (2018). Upregulácia Klotho prispieva k neuroprotekcii ligustilidu proti cerebrálnemu ischemickému poškodeniu u myší. Eur. J. Pharmacol. 820, 198-205. doi:10.1016/j.ejphar.2017.12.019
Luo, K. (2017). Krížová komunikácia medzi TGF-β/Smad a inými signálnymi dráhami. Cold Spring Harb. Perspektíva. Biol. 9 (1), a022137. doi:10.1101/cshperspect.a022137
Maekawa, Y., Ishikawa, K., Yasuda, O., Oguro, R., Hanasaki, H., Kida, I., et al. (2009). Klotho potláča TNF-a-indukovanú expresiu adhéznych molekúl v endoteli a tlmí aktiváciu NF-KB. Endocr 35 (3), 341–346. doi:10.1007/s12020-009-9181-3
Maizels, RM (2021). Mnohostranné úlohy TGF-β pri regulácii imunity voči infekcii. Adv. Immunol. 150, 1–42. doi:10.1016/bs.ai.2021.05.001
Maltese, G., Psefteli, P.-M., Rizzo, B., Srivastava, S., Gnudi, L., Mann, GE, et al. (2017). Hormón proti starnutiu klotho indukuje antioxidačnú obranu sprostredkovanú Nrf2 v bunkách hladkého svalstva ľudskej aorty. J. Cell. Mol. Med. 21 (3), 621-627. doi:10.1111/jcmm.12996
Manrique, C., Habibi, J., Aroor, AR, Sowers, JR, Jia, G., Hayden, MR, a kol. (2016). Inhibícia dipeptidylpeptidázy-4 linagliptínom zabraňuje vaskulárnym abnormalitám vyvolaným západnou stravou u samíc myší. Kardiovaskulárny diabetes. 15, 94. doi:10.1186/s12933-016-0414-5
Maquigussa, E., Paterno, JC, de Oliveira Pokorny, GH, da Silva Perez, M., Varela, VA, da Silva Novaes, A. a kol. (2018). Klotho a PPAR gama aktivácia sprostredkúvajú renoprotektívny účinok losartanu v modeli 5/6 nefrektómie. Predné. Physiol. 9, 1033. doi:10.3389/fphys.2018.01033
Maretzky, T., Scholz, F., Köten, B., Proksch, E., Saftig, P. a Reiss, K. (2008). ADAM10-sprostredkované uvoľňovanie E-kadherínu je regulované prozápalovými cytokínmi a moduluje súdržnosť keratinocytov pri ekzematóznej dermatitíde. J. Investigative Dermatology 128 (7), 1737–1746. doi:10.1038/sj.jid.5701242
Marsell, R., Krajisnik, T., Goransson, H., Ohlsson, C., Ljunggren, O., Larsson, TE, a kol. (2008). Analýza génovej expresie obličiek z transgénnych myší exprimujúcich fibroblastový rastový faktor-23. Nephrol. Vytáčať. Transplantácia. 23 (3), 827-833. doi:10.1093/ndt/gfm672
Martín-Núñez, E., Pérez-Castro, A., Tagua, VG, Hernández-Carballo, C., Ferri, C., Pérez-Delgado, N., et al. (2022). Expresia Klotho v periférnych krvných cirkulujúcich bunkách je spojená s vaskulárnym a systémovým zápalom pri aterosklerotickom vaskulárnom ochorení. Sci. Rep. 12 (1), 8422. doi:10.1038/s41598-022-12548-z
Mathew, R., Pal Bhadra, M. a Bhadra, U. (2017). Regulácia dĺžky života naprieč druhmi založená na signalizácii inzulínu/inzulínu podobného rastového faktora-1 (IIS). Biogerontológia 18 (1), 35–53. doi:10.1007/s10522-016-9670-8
Mehan, S., Bhalla, S., Siddiqui, EM, Sharma, N., Shandilya, A., and Khan, A. (2022). Potenciálne úlohy glukagónu podobného peptidu-1 a jeho analógov pri demencii zameranej na narušenú sekréciu inzulínu a neurodegeneráciu. Dnnd Vol. 12, 31-59. doi:10.2147/DNND.S247153
Mencke, R., Harms, G., Moser, J., van Meurs, M., Diepstra, A., Leuvenink, HG, a kol. (2017a). Ľudská alternatíva Klotho mRNA je nezmyselne sprostredkovaný cieľ rozpadu mRNA neefektívne zostrihaný pri ochorení obličiek. JCI Insight 2 (20), e94375. doi:10.1172/jci.insight.94375
Mencke, R., Olauson, H. a Hillebrands, J.-L. (2017b2017b). Účinky Klotho na fibrózu a rakovinu: Renálne zameranie na mechanizmy a terapeutické stratégie. Adv. Drug Deliv. 121 , 85–100. doi:10.1016/j.addr.2017.07.009
Mikula-Pietrasik, J., Rutecki, S. a Książek, K. (2022). Funkčná multipotencia signalizácie transformujúceho rastového faktora β na priesečníku starnutia a rakoviny. Cell. Mol. Life Sci. 79 (4), 196. doi:10.1007/s00018-022-04236-y
Mizusaki, K., Hasuike, Y., Kimura, T., Nagasawa, Y., Kuragano, T., Yamada, Y., et al. (2019). Inhibícia cicavčieho cieľa rapamycínu môže zvýšiť zvýšenie hladín rozpustného klotho u príjemcov transplantácie obličiek. Blood Purif. 47 (Suppl. 2), 12-18. doi:10.1159/000496630
Mohammed, I., Hollenberg, MD, Ding, H., a Triggle, ČR (2021). Kritický prehľad dôkazov, že metformín je domnelý liek proti starnutiu, ktorý zlepšuje zdravie a predlžuje životnosť. Predné. Endocrinol. 12, 718942. doi:10.3389/fendo.2021.718942
Moreira, PI (2014). Metformín v mozgu diabetika: priateľ alebo nepriateľ? Ann. Prekl. Med. 2 (6), 54. doi:10.3978/j.issn.2305-5839.2014.06.10
Moreno, JA, Izquierdo, MC, Sanchez-Niño, MD, Suárez-Alvarez, B., Lopez-Larrea, C., Jakubowski, A., et al. (2011). Zápalové cytokíny TWEAK a TNFα znižujú renálnu expresiu Klotho prostredníctvom NFκB. J Am Soc Nephrol 22 (7), 1315–1325. doi:10.1681/ASN.2010101073
Morishima, T. a Ochi, E. (2021). Vplyv jediného záchvatu odporového cvičenia na sérum Klotho u zdravých mladých mužov. Physiol. Rep. 9 (21), e15087. doi:10.14814/phy2.15087
Mullen, AC a Wrana, JL (2017). Signalizácia rodiny TGF-β pri obnove a diferenciácii embryonálnych a somatických kmeňových buniek. Cold Spring Harb. Perspektíva. Biol. 9 (7), a022186. doi:10.1101/cshperspect.a022186
Murphy, G. (2009). Regulácia proteolytických dezintegrínových metaloproteináz, ‚Sheddáz‘. Semináre Cell. & Dev. Biol. 20 (2), 138-145. doi:10.1016/j.semcdb.2008.09.004
Mýtych, J. (2022). Pôsobenie Klotho na hipokampálne neurónové bunky. Vitam. Horm. 118, 223-246. doi:10.1016/bs.vh.2021.12.001
Myung, J., Beom, J.-H., Kim, J.-H., Woo, J.-S., Park, I., Chung, S.-P., a kol. (2022). Rekombinantný proteín Klotho zlepšuje ischémiu myokardu/reperfúzne poškodenie zmiernením sterilného zápalu. Biomedicines 10 (4), 894. doi:10.3390/biomedicines10040894
Narumiya, H., Sasaki, S., Kuwahara, N., Irie, H., Kusaba, T., Kameyama, H., et al. (2004). Inhibítory HMG-CoA reduktázy up-regulujú klotho mRNA proti starnutiu prostredníctvom inaktivácie RhoA v bunkách IMCD3. Cardiovasc. Res. 64 (2), 331-336. doi:10.1016/j.cardiores.2004.07.011
Navarro-González, JF, Sánchez-Niño, MD, Donate-Correa, J., Martín-Núñez, E., Ferri, C., Pérez-Delgado, N., et al. (2018). Účinky pentoxifylínu na koncentrácie rozpustného Klotho a expresiu renálnych tubulárnych buniek pri diabetickej obličkovej chorobe. Diabetes Care 41 (8), 1817–1820. doi:10.2337/dc18-0078
Neitzel, J., Franzmeier, N., Rubinski, A., Dichgans, M., Brendel, M., au, fnm, et al. (2021). Heterozygotnosť KL-VS je spojená s nižšou akumuláciou tau závislou od amyloidu a poruchou pamäti pri Alzheimerovej chorobe. Nat. komun. 12 (1), 3825. doi:10.1038/s41467-021-23755-z
Neyra, JA, Hu, MC a Moe, OW (2021). Klotho v klinickej nefrológii. J Am Soc Nephrol 16 (1), 162–176. doi:10.2215/CJN.02840320
Neyra, JA, Moe, OW, Pastor, J., Gianella, F., Sidhu, SS, Sarnak, MJ, et al. (2020). Vykonávanie rozpustných testov Klotho v klinických vzorkách ochorenia obličiek. Clin. Obličky J. 13 (2), 235-244. doi:10.1093/ckj/sfz085
Ni, W., Zhang, Y. a Yin, Z. (2021). Ochranný mechanizmus mezenchymálnych kmeňových buniek kostnej drene modifikovaných génom Klotho pri akútnom poškodení obličiek vyvolanom rabdomyolýzou. Regen. Ther. 18, 255-267. doi:10.1016/j.reth.2021.07.003
Nie, F., Wu, D., Du, H., Yang, X., Yang, M., Pang, X., et al. (2017). Hladiny klotho proteínu v sére a ich korelácie s progresiou diabetes mellitus 2. typu. J. Diabetes jeho komplikácia. 31 (3), 594-598. doi:10.1016/j.jdiacomp.2016.11.008
Nilles, KL, Williams, EI, Betterton, RD, Davis, TP a Ronaldson, PT (2022). Transportéry hematoencefalickej bariéry: Príležitosti pre terapeutický rozvoj pri ischemickej cievnej mozgovej príhode. Ijms 23 (3), 1898. doi:10.3390/ijms23031898
Oh, HJ, Nam, BY, Wu, M., Kim, S., Park, J., Kang, S., a kol. (2018). Klotho hrá ochrannú úlohu proti glomerulárnej hypertrofii spôsobom závislým od bunkového cyklu pri diabetickej nefropatii. Am. J. Fyziológia-Fyziológia obličiek 315 (4), F791–F805. doi:10.1152/ajprenal.00462.2017
Ohrui, T., Matsui, T., Yamaya, M., Arai, H., Ebihara, S., Maruyama, M., et al. (2004). Inhibítory enzýmu konvertujúceho angiotenzín a výskyt Alzheimerovej choroby v Japonsku. J. Am. Geriatr. Soc. 52 (4), 649-650. doi:10.1111/j.1532-5415.2004.52178_7.x
Oishi, H., Doi, S., Nakashima, A., Ike, T., Maeoka, Y., Sasaki, K., et al. (2021). Nadmerná expresia Klotho chráni pred starnutím obličiek spolu s potlačením signálnych dráh transformujúceho rastového faktora-β1. Am. J. Fyziológia-Fyziológia obličiek 321 (6), F799–F811. doi:10.1152/ajprenal.00609.2020
Olejnik, A., Franczak, A., Krzywonos-Zawadzka, A., Kałużna-Oleksy, M., and Bil-Lula, I. (2018). Biologická úloha klotho proteínu vo vývoji kardiovaskulárnych chorôb. BioMed Res. Int. 2018, 1.–17. doi:10.1155/2018/5171945
Olejnik, A., Krzywonos‐Zawadzka, A., Banaszkiewicz, M. a Bil‐Lula, I. (2020). Klotho proteín prispieva ku kardioprotekcii počas ischémie/reperfúzneho poškodenia. J. Cell. Mol. Medi 24 (11), 6448-6458. doi:10.1111/jcmm.15293
Ostojič, SM a Engeset, D. (2021). Zlepšenie príjmu kreatínu v mozgu pomocou proteínovej stimulácie Klotho: môže diéta zasiahnuť? Predné. Nutr. 8, 795599. doi:10.3389/fnut.2021.795599
Palma, E., Ruffolo, G., Cifelli, P., Roseti, C., Vliet, EA v., a Aronica, E. (2018). Modulácia GABAA receptorov pri liečbe epilepsie. Cpd 23 (37), 5563-5568. doi:10.2174/1381612823666170809100230
Panda, H., Wen, H., Suzuki, M. a Yamamoto, M. (2022). Mnohostranné úlohy systému KEAP1-NRF2 v prostredí rakoviny a zápalových chorôb. Antioxidanty 11 (3), 538. doi:10.3390/antiox11030538
Pardridge, WM (2012). Transport liečiva cez hematoencefalickú bariéru. J. Cereb. Prietok krvi. Metab. 32 (11), 1959-1972. doi:10.1038/jcbfm.2012.126
Pawlak, JB a Blobe, GC (2022). Koreceptory TGF ‐β superrodiny pri rakovine. Dev. Dyn. 251 (1), 117–143. doi:10.1002/dvdy.338
Phan, P., Saikia, BB, Sonnaila, S., Agrawal, S., Alraawi, Z., Kumar, TKS, a kol. (2021). Sága endokrinných FGF. Bunky 10 (9), 2418. doi:10.3390/cells10092418
Pratsinis, H., Mavrogonatou, E. a Kletsas, D. (2017). „TGF-β vo vývoji a starnutí“, v Hormóny v starnutí a dlhovekosti . Gewerbestrasse 11, 6330 . Editor SIS Rattan (Cham, Švajčiarsko: Springer International Publishing AG ), 127–148. doi:10.1007/978-3-319-63001-4_7
Prud’homme, GJ (2012). Rakovinové kmeňové bunky a nové ciele pre protinádorové stratégie. Cpd 18 (19), 2838-2849. doi:10.2174/138161212800626120
Prud’homme, GJ, Glinka, Y., Kurt, M., Liu, W., and Wang, Q. (2017a). Proteín proti starnutiu Klotho je indukovaný terapiou GABA a má ochranné a stimulačné účinky na beta bunky pankreasu. Biochem. Biophysical Res. komun. 493 (4), 1542–1547. doi:10.1016/j.bbrc.2017.10.029
Prud’homme, GJ (2007). Patobiológia transformujúceho rastového faktora β pri rakovine, fibróze a imunologickom ochorení a terapeutické úvahy. Lab. Vyšetrovať. 87 (11), 1077-1091. doi:10.1038/labinvest.3700669
Prud’homme, GJ, Glinka, Y., Hasilo, C., Paraskevas, S., Li, X. a Wang, Q. (2013). GABA chráni bunky ľudských ostrovčekov pred škodlivými účinkami imunosupresívnych liekov a sama o sebe má imunoinhibičné účinky. Transplantácia 96 (7), 616–623. doi:10.1097/TP.0b013e31829c24be
Prud’homme, GJ, Glinka, Y., Kurt, M., Liu, W. a Wang, Q. (2020). Systémová terapia Klotho chráni pred inzulitídou a zvyšuje hmotnosť beta-buniek u NOD myší. Biochem. Biophysical Res. komun. 525 (3), 693-698. doi:10.1016/j.bbrc.2020.02.123
Prud’homme, GJ, Glinka, Y., Matkar, PN, a Leong-Poi, H. (2017b). „Úloha neuropilínov v signalizácii TGF-β a biológii rakoviny“, v knihe Neuropilins: Role and Function in Health and Disease . Editori G. Neufeld a O. Kessler (Cham: Sprinter International Publishing AG ), 187–212. doi:10.1007/978-3-319-48824-0_11
Prud’homme, GJ a Glinka, Y. (2012). Neuropilíny sú multifunkčné koreceptory, ktoré sa podieľajú na iniciácii, raste, metastázovaní a imunite nádoru. Oncotarget 3 (9), 921-939. doi:10.18632/oncotarget.626
Pujadas, G. a Drucker, DJ (2016). Vaskulárna biológia peptidov superrodiny glukagónových receptorov: mechanický a klinický význam. Endokr. 37 (6), 554-583. doi:10.1210/er.2016-1078
Purwana, I., Zheng, J., Li, X., Deurloo, M., Son, DO, Zhang, Z., a kol. (2014). GABA podporuje proliferáciu ľudských β-buniek a moduluje homeostázu glukózy. Diabetes 63 (12), 4197-4205. doi:10.2337/db14-0153
Qu, Z., Sun, J., Zhang, W., Yu, J. a Zhuang, C. (2020). Transkripčný faktor NRF2 ako sľubný terapeutický cieľ pre Alzheimerovu chorobu. Voľný Radic. Biol. Med. 159, 87-102. doi:10.1016/j.freeradbiomed.2020.06.028
Rao, YL, Ganaraja, B., Murlimanju, BV, Joy, T., Krishnamurthy, A., and Agrawal, A. (2022). Hippokampus a jeho zapojenie do Alzheimerovej choroby: prehľad. 3 Biotech. 12 (2), 55. doi:10.1007/s13205-022-03123-4
Rao, Z., Landry, T., Li, P., Bunner, W., Laing, BT, Yuan, Y., a kol. (2019). Podávanie alfa klotho znižuje akumuláciu pečeňových a tukových lipidov u obéznych myší. Heliyon 5 (4), e01494. doi:10.1016/j.heliyon.2019.e01494
Rausch, S. a Föller, M. (2022). Regulácia FGF23 za fyziologických a patofyziologických podmienok. Pflugers Arch. – Eur. J. Physiol. 474 (3), 281–292. doi:10.1007/s00424-022-02668-w
Razzaque, MS (2012). Úloha Klotho v energetickom metabolizme. Nat. Endocrinol. 8 (10), 579-587. doi:10.1038/nrendo.2012.75
Reddy, PH, Manczak, M., Yin, X., Grady, MC, Mitchell, A., Tonk, S., a kol. (2018). Ochranné účinky indického korenia kurkumínu proti amyloidu-β pri Alzheimerovej chorobe. Jad 61 (3), 843-866. doi:10.3233/JAD-170512
Rim, EY, Clevers, H. a Nusse, R. (2022). Cesta Wnt: Od signalizačných mechanizmov k syntetickým modulátorom. Annu. Biochem. 91. doi:10.1146/annurev-biochem-040320-103615
Roberti, A., Chaffey, LE a Greaves, DR (2022). NF-KB signalizácia a zápal – premena lieku na liečbu zápalových porúch? Biológia 11 (3), 372. doi:10.3390/biology11030372
Rubinek, T. a Wolf, I. (2016). Úloha Alpha-Klotho ako univerzálneho supresora nádorov. Vitam. Horm. 101, 197-214. doi:10.1016/bs.vh.2016.03.001
Ruoslahti, E. (2017). Peptidy prenikajúce do nádoru na zlepšenie dodávania liečiva. Adv. Drug Deliv. Zj 110-111, 3-12. doi:10.1016/j.addr.2016.03.008
Saar-Kovrov, V., Donners, MMPC a van der Vorst, EPC (2021). Vylučovanie Klotho: Funkčné dôsledky pri chronickom ochorení obličiek a pridružených vaskulárnych ochoreniach. Predné. Cardiovasc. Med. 7, 617842. doi:10.3389/fcvm.2020.617842
Sachdeva, A., Gouge, J., Kontovounisios, C., Nikolaou, S., Ashworth, A., Lim, K., et al. (2020). Klotho a liečba ľudských malignít. Cancers 12 (6), 1665. doi:10.3390/cancers12061665
Sanz, B., Arrieta, H., Rezola-Pardo, C., Fernández-Atutxa, A., Garin-Balerdi, J., Arizaga, N., et al. (2021). Nízka koncentrácia klotho v sére je spojená s horším poznávaním, psychickými komponentmi krehkosti, závislosti a pádmi u obyvateľov domovov dôchodcov. Sci. Rep. 11 (1), 9098. doi:10.1038/s41598-021-88455-6
Sarang, SS, Lukyanova, SM, Brown, DD, Cummings, BS, Gullans, SR a Schnellmann, RG (2008). Identifikácia, spoluzostavenie a aktivita podjednotiek receptora kyseliny y-aminomaslovej v renálnych proximálnych tubulárnych bunkách. J. Pharmacol. Exp. Ther. 324 (1), 376–382. doi:10.1124/jpet.107.129957
Sarnobat, D., Charlotte Moffett, R., Flatt, PR, Irwin, N. a Tarasov, AI (2022). GABA a inzulín, ale nie nikotínamid, zvyšujú transdiferenciáciu α- na β-buniek u diabetických myší s deficitom inzulínu. Biochem. Pharmacol. 199, 115019. doi:10.1016/j.bcp.2022.115019
Sasaki, S., Tohda, C., Kim, M. a Yokozawa, T. (2007). Kyselina gama-aminomaslová špecificky inhibuje progresiu tubulárnej fibrózy a atrofie u nefrektomizovaných potkanov. Biol. Pharm. Bull. 30 (4), 687-691. doi:10.1248/bpb.30.687
Semba, RD, Moghekar, AR, Hu, J., Sun, K., Turner, R., Ferrucci, L., et al. (2014). Klotho v cerebrospinálnej tekutine dospelých s Alzheimerovou chorobou a bez nej. Neurosci. Lett. 558, 37–40. doi:10.1016/j.neulet.2013.10.058
Shardell, M., Semba, RD, Rosano, C., Kalyani, RR, Bandinelli, S., Chia, CW, a kol. (2016). Plazmové klotho a kognitívny pokles u starších dospelých: zistenia zo štúdie InCHIANTI. Gerona 71 (5), 677–682. doi:10.1093/gerona/glv140
Shen, D., Li, H., Zhou, R., Liu, M.-j., Yu, H. a Wu, D.-F. (2018). Pioglitazón zmierňuje poruchy súvisiace so starnutím u starých myší s deficitom apolipoproteínu E. Exp. Gerontol. 102, 101-108. doi:10.1016/j.exger.2017.12.002
Shin, YJ, Luo, K., Quan, Y., Ko, EJ, Chung, BH, Lim, SW, a kol. (2019). Terapeutická výzva minikruhového vektorového kódovania Klotho vo zvieracom modeli. Am. J. Nephrol. 49 (5), 413-424. doi:10.1159/000499863
Soltani, N., Qiu, H., Aleksic, M., Glinka, Y., Zhao, F., Liu, R., et al. (2011). GABA má ochranné a regeneračné účinky na beta bunky ostrovčekov a zvráti cukrovku. Proc. Natl. Akad. Sci. USA 108 (28), 11692–11697. doi:10.1073/pnas.1102715108
Son, DO, Liu, W., Li, X., Prud’homme, GJ a Wang, Q. (2019). Kombinovaný účinok GABA a agonistu receptora glukagónu podobného peptidu-1 na apoptózu indukovanú cytokínom v pankreatickej β-bunkovej línii a izolovaných ľudských ostrovčekoch. J. Diabetes 11 (7), 563-572. doi:10.1111/1753-0407.12881
Soo, SK, Rudich, PD, Traa, A., Harris-Gauthier, N., Shields, HJ a Van Raamsdonk, JM (2020). Zlúčeniny, ktoré predlžujú dlhovekosť, sú ochranné pri neurodegeneratívnych ochoreniach a poskytujú novú liečebnú stratégiu pre tieto devastujúce poruchy. Mech. Starnutie Dev. 190, 111297. doi:10.1016/j.mad.2020.111297
Sosa-Pineda, B., Chowdhury, K., Torres, M., Oliver, G. a Gruss, P. (1997). Gén Pax4 je nevyhnutný pre diferenciáciu β buniek produkujúcich inzulín v pankrease cicavcov. Príroda 386 (6623), 399–402. doi:10.1038/386399a0
Tabibzadeh, S. (2021). Signálne dráhy a efektory starnutia. Predné. Biosci. 26 (1), 50-96. doi:10.2741/4889
Takano, K., Yatabe, MS, Abe, A., Suzuki, Y., Sanada, H., Watanabe, T., a kol. (2014). Charakteristické prejavy GABA receptorov a molekúl produkujúcich/transportujúcich GABA v obličkách potkanov. PLoS One 9 (9), e105835. doi:10.1371/journal.pone.0105835
Takenaka, T., Inoue, T., Miyazaki, T., Kobori, H., Nishiyama, A., Ishii, N., et al. (2018). Klotho zlepšuje medulárnu fibrózu a tlakovú natriurézu u hypertenzných obličiek potkanov. Hypertenzia 72 (5), 1151–1159. doi:10.1161/HYPERTENSIONAHA.118.11176
Takenaka, T., Inoue, T., Miyazaki, T., Kobori, H., Nishiyama, A., Ishii, N., et al. (2017). Klotho potláča renín-angiotenzínový systém pri adriamycínovej nefropatii. Nephrol. Vytáčať. Transpl. 32 (5), gfw340–800. doi:10.1093/ndt/gfw340
Takenaka, T., Kobori, H., Inoue, T., Miyazaki, T., Suzuki, H., Nishiyama, A., et al. (2020). Suplementácia Klotho zlepšuje krvný tlak a funkciu obličiek u myší DBA/2-pcy, čo je model polycystickej choroby obličiek. Am. J. Fyziológia-Fyziológia obličiek 318 (3), F557–F564. doi:10.1152/ajprenal.00299.2019
Takenaka, T., Kobori, H., Miyazaki, T., Suzuki, H., Nishiyama, A., Ishii, N., et al. (2019). Suplementácia proteínu Klotho znižuje krvný tlak a renálnu hypertrofiu u db/db myší, čo je model diabetu 2. Acta Physiol. 225 (2), e13190. doi:10.1111/apha.13190
Takeshita, A., Kawakami, K., Furushima, K., Miyajima, M. a Sakaguchi, K. (2018). Centrálna úloha proximálneho tubulárneho aKlotho/FGF receptorového komplexu v metabolizme fosfátu a vitamínu D regulovanom FGF23. Sci. Rep. 8 (1), 6917. doi:10.1038/s41598-018-25087-3
Tan, S.-J., Chu, MM, Toussaint, ND, Cai, MM, Hewitson, TD a Holt, SG (2018). Vysoko intenzívne fyzické cvičenie zvyšuje sérové hladiny α-klotho u zdravých dobrovoľníkov. J. Circulating Biomarkers 7, 184945441879458. doi:10.1177/1849454418794582
Tarhani, F., Heidari, G. a Nezami, A. (2020). Hodnotenie hladiny α-klotho u detí s inzulín-dependentným diabetes mellitus (IDDM). J. Pediatr. Endocrinol. Metab. 33 (6), 761-765. doi:10.1515/jpem-2019-0591
Tasnim, N., Dutta, P., Nayeem, J., Masud, P., Ferdousi, A., Ghosh, AS, a kol. (2021). Osteoporóza, nevyhnutná okolnosť chronického ochorenia obličiek: Systematický prehľad. Cureus 13 (10), e18488. doi:10.7759/cureus.18488
Tataranni, T., Biondi, G., Cariello, M., Mangino, M., Colucci, G., Rutigliano, M., et al. (2011). Rapamycínom indukovaná hypofosfatémia a inzulínová rezistencia sú spojené s aktiváciou mTORC2 a expresiou Klotho. Am. J. Transpl. 11 (8), 1656–1664. doi:10.1111/j.1600-6143.2011.03590.x
Thal, DR, von Arnim, C., Griffin, WST, Yamaguchi, H., Mrak, RE, Attems, J., et al. (2013). Patológia klinickej a predklinickej Alzheimerovej choroby. Eur. Arch. Psychiatry Clin. Neurosci. 263 (Suppl. 2), 137–145. doi:10.1007/s00406-013-0449-5
Tian, J., Dang, H., Chen, Z., Guan, A., Jin, Y., Atkinson, MA a kol. (2013). Kyselina γ-aminomaslová reguluje prežitie aj replikáciu ľudských β-buniek. Diabetes 62 (11), 3760–3765. doi:10.2337/db13-0931
Tian, J., Lu, Y., Zhang, H., Chau, CH, Dang, HN, a Kaufman, DL (2004). Kyselina y-aminomaslová inhibuje autoimunitu T-buniek a rozvoj zápalových odpovedí v myšom modeli diabetu 1. typu. J. Immunol. 173 (8), 5298-5304. doi:10.4049/jimmunol.173.8.5298
Tominaga, K. a Suzuki, HI (2019). Signalizácia TGF-β v bunkovom starnutí a patológii súvisiacej so starnutím. Ijms 20 (20), 5002. doi:10.3390/ijms20205002
Tsujikawa, H., Kurotaki, Y., Fujimori, T., Fukuda, K. a Nabeshima, Y.-I. (2003). Klotho, gén súvisiaci so syndrómom pripomínajúcim ľudské predčasné starnutie, funguje v negatívnom regulačnom okruhu endokrinného systému vitamínu D. Mol. Endocrinol. 17 (12), 2393-2403. doi:10.1210/me.2003-0048
Typiak, M. a Piwkowska, A. (2021). Protizápalové účinky Klotho: Dôsledky pre terapiu diabetickej nefropatie. Ijms 22 (2), 956. doi:10.3390/ijms22020956
Tyurenkov, IN, Perfilova, VN, Nesterova, AA a Glinka, Y. (2021). Klotho proteín a kardiovaskulárny systém. Biochem. Mosc. 86 (2), 132–145. doi:10.1134/S0006297921020024
Untereiner, A., Abdo, S., Bhattacharjee, A., Gohil, H., Pourasgari, F., Ibeh, N., a kol. (2019). GABA podporuje proliferáciu β-buniek, ale neprekonáva zhoršenú glukózovú homeostázu spojenú s obezitou vyvolanou diétou. FASEB J. 33 (3), 3968-3984. doi:10.1096/fj.201801397R
Urakawa, I., Yamazaki, Y., Shimada, T., Iijima, K., Hasegawa, H., Okawa, K., et al. (2006). Klotho konvertuje kanonický receptor FGF na špecifický receptor pre FGF23. Príroda 444 (7120), 770–774. doi:10.1038/nature05315
van Loon, EPM, Pulskens, WP, van der Hagen, EAE, Lavrijsen, M., Vervloet, MG, van Goor, H. a kol. (2015). Vylučovanie klotho prostredníctvom ADAMov v obličkách. Am. J. Fyziológia-Fyziológia obličiek 309 (4), F359–F368. doi:10.1152/ajprenal.00240.2014
Veronesi, F., Borsari, V., Cherubini, A. a Fini, M. (2021). Asociácia Klotho s fyzickým výkonom a krehkosťou u dospelých v strednom veku a starších: Systematický prehľad. Exp. Gerontol. 154, 111518. doi:10.1016/j.exger.2021.111518
Vo, HT, Laszczyk, AM a King, GD (2018). Klotho, kľúč k zdravému starnutiu mozgu? Bpl 3 (2), 183-194. doi:10.3233/BPL-170057
Wan, Q., He, Y. a Yuan, M. (2017). Klotho pri cukrovke a diabetickej nefropatii: stručný prehľad aktualizácie. Int. J. Clin. Exp. Med. 10 (3), 4342-4349. Dostupné na: www.ijcem.com/ISSN:1940-5901/IJCEM0044613 .
Wang, F. a Zheng, J. (2022). Súvislosť medzi sérovým alfa-Klotho a ťažkou kalcifikáciou brušnej aorty medzi civilistami v Spojených štátoch. Nutr. Metabolizmus Cardiovasc. Dis. 32 (22), 00106-01492. doi:10.1016/j.numecd.2022.02.017
Wang, Q., Prud’homme, G. a Wan, Y. (2015). GABAergický systém v endokrinnom pankrease: nový cieľ liečby cukrovky. Dmso 8, 79–87. doi:10.2147/DMSO.S50642
Wang, Q., Ren, L., Wan, Y. a Prud’homme, GJ (2019). GABAergická regulácia buniek pankreatických ostrovčekov: Fyziológia a antidiabetické účinky. J. Cell. Fyziológia 234 (9), 14432–14444. doi:10.1002/jcp.28214
Wang, Y. a Sun, Z. (2009). Súčasné chápanie klotho. Ageing Res. 8 (1), 43–51. doi:10.1016/j.arr.2008.10.002
Wolf, I., Levanon-Cohen, S., Bose, S., Ligumsky, H., Sredni, B., Kanety, H., et al. (2008). Klotho: supresor nádoru a modulátor dráh IGF-1 a FGF pri ľudskej rakovine prsníka. Oncogene 27 (56), 7094-7105. doi:10.1038/onc.2008.292
Wood, WG, Eckert, GP, Igbavboa, U., a Müller, WE (2010). Statíny a neuroprotekcia: Predpis na posunutie poľa vpred. Ann. NY Acad. Sci. 1199, 69-76. doi:10.1111/j.1749-6632.2009.05359.x
Wu, S.-E. a Chen, W.-L. (2022). Rozpustné klotho ako účinný biomarker na charakterizáciu zápalových stavov. Ann. Med. 54 (1), 1520–1529. doi:10.1080/07853890.2022.2077428
Xiang, T., Luo, X., Ye, L., Huang, H. a Wu, Y. (2022). Klotho zmierňuje neurozápal sprostredkovaný zápalom NLRP3 u potkanieho modelu epilepsie temporálneho laloku aktiváciou signálnej dráhy Nrf2. Epilepsia a správanie. 128, 108509. doi:10.1016/j.yebeh.2021.108509
Xiang, T., Luo, X., Zeng, C., Li, S., Ma, M. a Wu, Y. (2021). Klotho zlepšil kognitívne deficity v modeli potkanov s epilepsiou temporálneho laloku inhibíciou ferroptózy. Brain Res. 1772, 147668. doi:10.1016/j.brainres.2021.147668
Xie, B., Zhou, J., Shu, G., Liu, D.-c., Zhou, J., Chen, J., et al. (2013). Obnova expresie génu klotho indukuje apoptózu a autofágiu v bunkách rakoviny žalúdka: supresívna úloha klotho pri rakovine žalúdka. Cancer Cell. Int. 13 (1), 18. doi:10.1186/1475-2867-13-18
Xing, L., Guo, H., Meng, S., Zhu, B., Fang, J., Huang, J., et al. (2021). Klotho zlepšuje diabetickú nefropatiu aktiváciou signálnej dráhy Nrf2 v podocytoch. Biochem. Biophysical Res. komun. 534, 450-456. doi:10.1016/j.bbrc.2020.11.061
Xu, Y. a Sun, Z. (2015). Molekulárny základ Klotho: od génu k funkcii pri starnutí. Endokr. 36 (2), 174–193. doi:10.1210/er.2013-1079
Xue, J., Wang, L., Sun, Z. a Xing, C. (2019). Základný výskum v zdravotnej starostlivosti o diabetickú nefropatiu: Štúdia renoprotektívneho mechanizmu metformínu. J. Med. Syst. 43 (8), 266. doi:10.1007/s10916-019-1412-4
Yamamoto, M., Clark, JD, Pastor, JV, Gurnani, P., Nandi, A., Kurosu, H., a kol. (2005). Regulácia oxidačného stresu hormónom proti starnutiu klotho. J. Biol. Chem. 280 (45), 38029–38034. doi:10.1074/jbc.M509039200
Yang, K., Nie, L., Huang, Y., Zhang, J., Xiao, T., Guan, X., et al. (2012). Zlepšenie dysfunkcie endotelových buniek indukovanej uremickým toxínom indoxylsulfátom pomocou proteínu Klotho. Toxicol. Lett. 215 (2), 77-83. doi:10.1016/j.toxlet.2012.10.004
Yang, K., Wang, C., Nie, L., Zhao, X., Gu, J., Guan, X. a kol. (2015). Zhao J Klotho chráni pred hypertrofiou myokardu vyvolanou indoxylsulfátom. J. Am. Soc. Nephrol. 26 (10), 2434-2446. doi:10.1681/ASN.2014060543
Yoon, HE, Ghee, JY, Piao, S., Song, JH, Han, DH, Kim, S. a kol. (2011). Blokáda angiotenzínu II upreguluje expresiu Klotho, génu proti starnutiu, v experimentálnom modeli chronickej cyklosporínovej nefropatie. Nephrol. Vytáčať. Transpl. 26 (3), 800-813. doi:10.1093/ndt/gfq537
Yossef, RR, Al-Yamany, MF, Saad, MA a El-Sahar, AE (2020). Neuroprotektívne účinky vildagliptínu na liekmi indukovanú Alzheimerovu chorobu u potkanov s metabolickým syndrómom: Úloha Hippocampal Klotho a AKT signálnych dráh. Eur. J. Pharmacol. 889, 173612. doi:10.1016/j.ejphar.2020.173612
Yoon, HE, Lim, SW, Piao, SG, Song, JH, Kim, J. a Yang, CW (2012). Statín upreguluje expresiu klotho, génu proti starnutiu, pri experimentálnej cyklosporínovej nefropatii. Nephron Exp. Nephrol. 120 (4), e123–33. doi:10.1159/000342117
Yuan, Q., Ren, Q., Li, L., Tan, H., Lu, M., Tian, Y., a kol. (2022). Peptid odvodený od Klotho chráni pred fibrózou obličiek zacielením na signalizáciu TGF-beta. Nat. komun. 13 (1), 438. doi:10.1038/s41467-022-28096-z
Zeldich, E., Chen, CD, Colvin, TA, Bove-Fenderson, EA, Liang, J., Tucker Zhou, TB, a kol. (2014). Neuroprotektívny účinok Klotho je sprostredkovaný reguláciou členov redoxného systému. J. Biol. Chem. 289 (35), 24700–24715. [. doi:10.1074/jbc.M114.567321
Zeng, CY, Yang, TT, Zhou, HJ, Zhao, Y., Kuang, X., Duan, W. a kol. (2019). Nadmerná expresia Klotho v mozgu sprostredkovaná lentivírusovým vektorom zlepšuje patológiu podobnú Alzheimerovej chorobe a kognitívne deficity u myší. Neurobiol. Vek 78, 18–28. doi:10.1016/j.neurobiolaging.2019.02.003
Zhang, H., Li, Y., Fan, Y., Wu, J., Zhao, B., Guan, Y., et al. (2008). Klotho je cieľový gén PPAR-gama. Kidney Int. 74 (6), 732-739. doi:10.1038/ki.2008.244
Zhang, L. a Liu, T. (2018). Klinické dôsledky zmien sérových hladín Klotho u pacientov s diabetes mellitus 2. typu a s ním spojené komplikácie. J. Diabetes Complicat. 32 (10), 922-930. doi:10.1016/j.jdiacomp.2018.06.002
Zhang, P., Li, Y., Du, Y., Li, G., Wang, L. a Zhou, F. (2016). Resveratrol zlepšil vaskulárnu kalcifikáciu reguláciou Sirt-1 a Nrf2. Transpl. Proc. 48 (10), 3378-3386. doi:10.1016/j.transproceed.2016.10.023
Zhang, T., Ma, C., Zhang, Z., Zhang, H. a Hu, H. (2020). Signalizácia NF-kappaB pri zápale a rakovine. MedComm 2 (4), 618-653. doi:10.1002/mco2.104
Zhang, XT, Wang, G., Ye, LF, Pu, Y., Li, RT, Liang, J. a kol. (2020). Baicalinová reverzia supresie Klotho spojenej s hypermetyláciou DNA zlepšuje poškodenie obličiek v modeli diabetickej myši typu 1. Cell. Cyklus 19 (23), 3329–3347. doi:10.1080/15384101.2020.1843815
Zhao, L., Chen, H., Lu, L., Zhao, C., Malichewe, CV, Wang, L., et al. (2021). Návrh a skríning nového penetračného peptidu zacieleného na neuropilín-1 na antiangiogénnu terapiu pri glióme. Life Sci. 270, 119113. doi:10.1016/j.lfs.2021.119113
Zhao, Y., Banerjee, S., Dey, N., LeJeune, WS, Sarkar, PS, Brobey, R., a kol. (2011). Deplécia Klotho prispieva k zvýšenému zápalu v obličkách db/db myšacieho modelu diabetu prostredníctvom fosforylácie RelA (serín)536. Diabetes 60 (7), 1907–1916. doi:10.2337/db10-1262
Zhao, Y., Zhao, MM, Cai, Y., Zheng, MF, Sun, WL, Zhang, SY, a kol. (2015). Cicavčí cieľ inhibície signalizácie rapamycínu zlepšuje vaskulárnu kalcifikáciu prostredníctvom regulácie Klotho. Kidney Int. 88 (4), 711-721. doi:10.1038/ki.2015.160
Zhao, YN, Shao, X., Ouyang, LF, Chen, L. a Gu, L. (2018). Kvalitatívna detekcia ginsenozidov v mozgových tkanivách po perorálnom podaní vysoko čistých celkových saponínov ženšenu pomocou polyklonálnej protilátky proti ginsenosidom. Chin. J. Nat. Med. 16 (3), 175-183. doi:10.1016/S1875-5364(18)30045-1
Zhong, X., Jagarlapudi, S., Weng, Y., Ly, M., Rouse, JC, McClure, K., a kol. (2020). Vzťahy medzi štruktúrou a funkciou rozpustnej formy proteínu proti starnutiu Klotho majú terapeutické dôsledky na zvládnutie ochorenia obličiek. J. Biol. Chem. 295 (10), 3115-3133. doi:10.1074/jbc.RA119.012144
Zhou, Q., Lin, S., Tang, R., Veeraragoo, P., Peng, W. a Wu, R. (2010). Úloha fosinoprilu a valsartanu na expresiu génu Klotho indukovanej angiotenzínom II v renálnych tubulárnych epiteliálnych bunkách potkanov. Kidney Blood Press Res. 33 (3), 186–192. doi:10.1159/000316703
Zhou, W., Chen, MM, Liu, HL, Si, ZL, Wu, WH, Jiang, H. a kol. (2022). Dihydroartemisinín potláča renálnu fibrózu u myší inhibíciou DNA-metyltransferázy 1 a zvýšením Klotho. Acta Pharmacol. Hriech . doi:10.1038/s41401-022-00898-3
Zhou, X., Yang, Q., Xie, Y., Sun, J., Hu, J., Qiu, P., et al. (2015). Tetrahydroxystilbénglukozid predlžuje životnosť myší prostredníctvom upregulácie nervového klotho a downregulácie nervového inzulínu alebo inzulínu podobného rastového faktora 1. Neurobiol. Starnutie 36 (3), 1462–1470. doi:10.1016/j.neurobiolaging.2014.11.002
Zhu, H., Gao, Y., Zhu, S., Cui, Q. a Du, J. (2017). Klotho zlepšuje srdcovú funkciu potlačením apoptózy sprostredkovanej reaktívnymi druhmi kyslíka (ROS) moduláciou signalizácie Mapks/Nrf2 pri kardiotoxicite vyvolanej doxorubicínom. Med. Sci. Monit. 23, 5283-5293. doi:10.12659/msm.907449
Zou, D., Wu, W., He, Y., Ma, S. a Gao, J. (2018). Úloha klotho pri chronickom ochorení obličiek. BMC Nephrol. 19 (1), 285. doi:10.1186/s12882-018-1094-z
Kľúčové slová: starnutie, FGF23, hyperfosfatémia, klotho, IGF-1, NF-KappaB, TGF-beta, Wnt
Citácia: Prud’homme GJ, Kurt M a Wang Q (2022) Pathobiology of the Klotho Antiaging Protein and Therapeutic Benefits. Predné. Starnutie 3:931331. doi: 10.3389/fragi.2022.931331
Prijaté: 28. apríla 2022; Prijaté: 6. júna 2022;
Zverejnené: 12. júla 2022.
Upravil:
Cátia F. Lourenço , Univerzita v Coimbre, Portugalsko
Skontroloval:
Mujib Ullah , Stanford University, Spojené štáty
Taylor Landry , East Carolina University, Spojené štáty americké
Copyright © 2022 Prud’homme, Kurt a Wang. Toto je článok s otvoreným prístupom distribuovaný v súlade s podmienkami licencie Creative Commons Attribution License (CC BY). Použitie, distribúcia alebo reprodukcia na iných fórach je povolená za predpokladu, že sú uvedení pôvodní autori a vlastník autorských práv a že sa cituje pôvodná publikácia v tomto časopise v súlade s uznávanou akademickou praxou. Nie je povolené žiadne použitie, distribúcia alebo reprodukcia, ktorá nie je v súlade s týmito podmienkami.
*Koešpondencia: Gérald J. Prud’homme, prudhommegj@gmail.com
Zrieknutie sa zodpovednosti: Všetky tvrdenia vyjadrené v tomto článku sú výlučne tvrdeniami autorov a nemusia nevyhnutne predstavovať tvrdenia ich pridružených organizácií alebo tvrdenia vydavateľa, redaktorov a recenzentov. Žiadny produkt, ktorý môže byť hodnotený v tomto článku, alebo tvrdenie, ktoré môže uviesť jeho výrobca, nie je zaručené ani schválené vydavateľom.



